Download presentation
Presentation is loading. Please wait.

1
Berg • Tymoczko • Stryer
Biochemistry Sixth Edition Chapter 7 Hemoglobin: Portrait of a Protein in Action Copyright © 2007 by W. H. Freeman and Company

5
Introduction to Heme Proteins
Biomedical importance Structure of heme Structure and function of Mb Structure and function of Hb O2 saturation curve of Mb and Hb differences The Bohr effect Effect of 2, 3 Bisphosphoglycerate Hemoglobinopathies Met Hb HbS and thalassemias

6
Biomedical Importance
Heme proteins function O2 binding O2 transport Electron transport Sickle cell disease: subunit of Hb altered. Cyanide and CO poisoning 2, 3 BPG (Bisphosphoglycerate) effect on Hb.
 effect on Hb.")
8
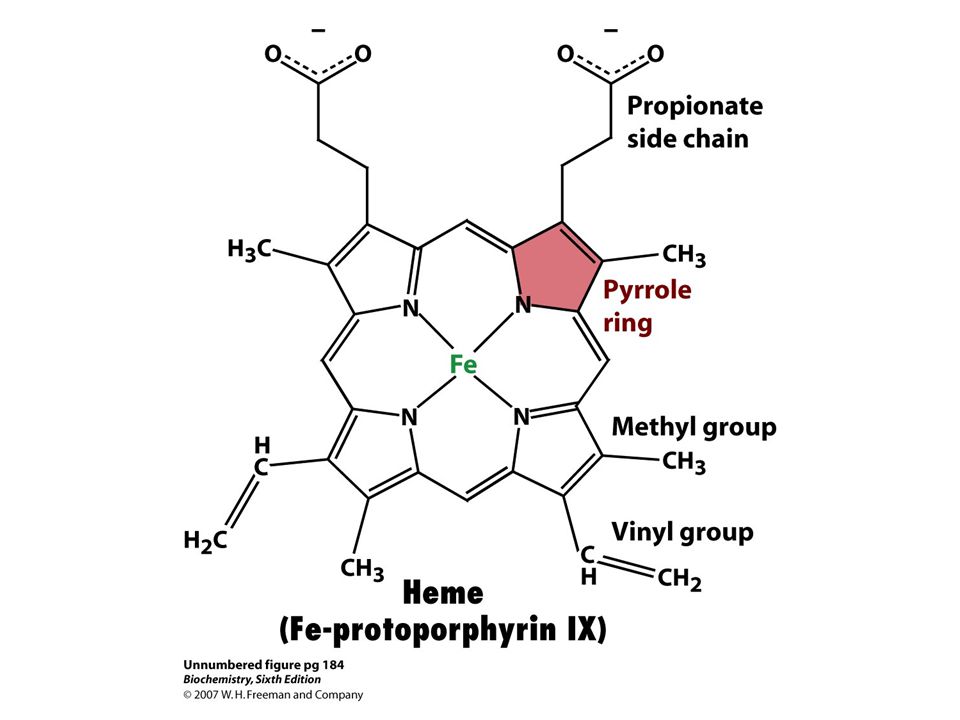
Heme Proteins Mb and Hb are “heme” proteins. Porphyrins are colored.
Iron porphyrin gives red color to blood. Magnesium porphyrin gives green color to plants. Fe2+ + protoporphyrin IX = HEME Heme is a prosthetic group, or non-polypeptide unit necessary for the biological activity. FerriMb-Iron is +3 oxidation state OxyMb-Iron is +2 oxidation state and O2 is bound to 6th DeoxyMb-Iron is +2 oxidation state

9
Heme Structure Four pyrolles linked methene bridges
Four CH3, 2 vinyl, and 2 propionate side chains could be arranged in fifteen different ways. Only one isomer, protoporphyrin IX, present. Fe2+ Four nitrogen atoms Fe2+ normally octahedrally coordinated. 6 ligands 4N of the porphyrin ring available 2 remaining In cytochrome c 5th and 6th met and His In Mb and Hb 5th coordinated to a His, 6th is for O2

10
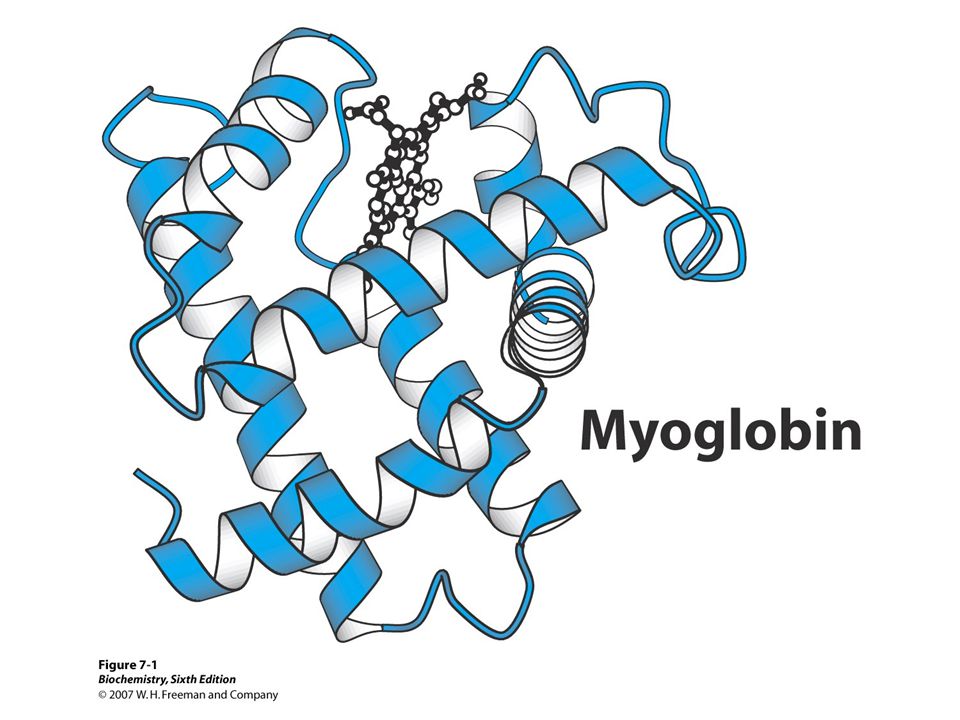
Structure and Function of Myoglobin
Myoglobin, a heme protein in heart and skeletal muscles. Myboglobin functions as reservoir for O2. carrier for O2. Myoglobin is single polypeptide chain (MW 17,000), 153 amino acids Surface is polar, interior is non-polar (characteristic pattern of globular proteins). 75% of polypeptide chain folded into eight stretches of -helical regions labeled A-H. Four segments terminated by proline.
, 153 amino acids. Surface is polar, interior is non-polar (characteristic pattern of globular proteins). 75% of polypeptide chain folded into eight stretches of -helical regions labeled A-H. Four segments terminated by proline.")
11
Structure and Function Continued
Heme group of Mb in crevice in proteins (between E and F helices). Cavity lined with non-polar amino acids, except two His residues. Proximal His binds Fe2+. Distal His (E7) does not directly interact with heme group but helps stabilize ferrous form of iron. Therefore, Mb creates special environment for heme that allows reversible binding of oxygen without simultaneous oxidation of Fe2+. His F8 means 8th residue in F helix, it identifies it as a histidyl residue. F8 proximal His E7 distal His
. Cavity lined with non-polar amino acids, except two His residues. Proximal His binds Fe2+. Distal His (E7) does not directly interact with heme group but helps stabilize ferrous form of iron. Therefore, Mb creates special environment for heme that allows reversible binding of oxygen without simultaneous oxidation of Fe2+. His F8 means 8th residue in F helix, it identifies it as a histidyl residue. F8 proximal His. E7 distal His.")
12
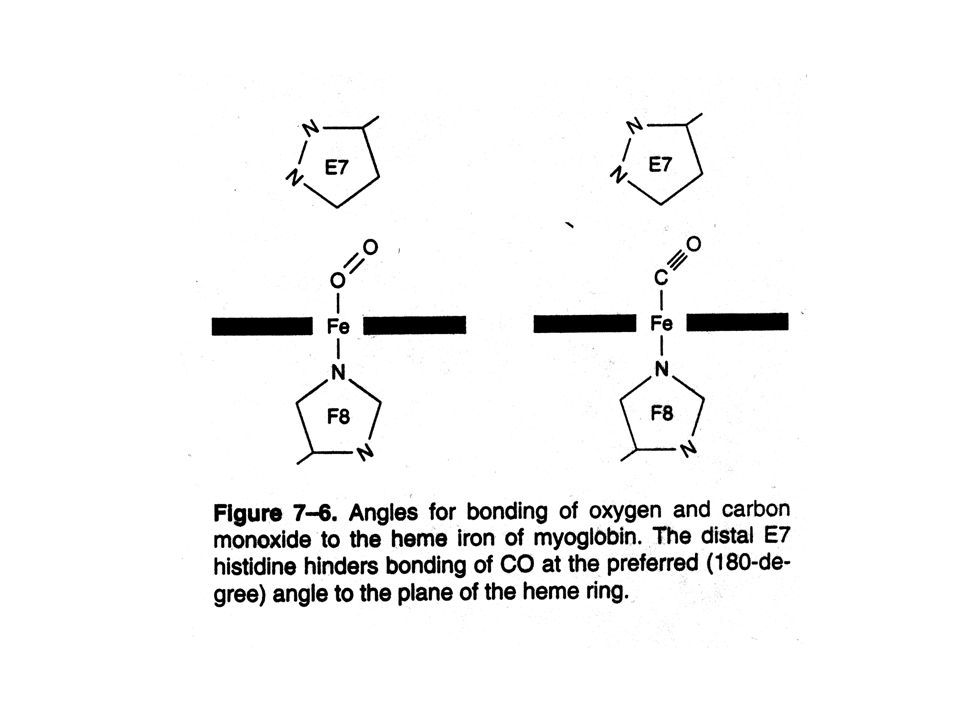
More Structure and Function
CO binds 25,000 times stronger than O2 in isolated heme. Because no distal His in isolated heme. In biological systems, distal His forces CO to bind at an angle, decreasing affinity of CO to heme (200 times). This protects us against CO poisoning. Natural protection.
. This protects us against CO poisoning. Natural protection.")
13
Electron-density Map of Myoglobin Near the Oxygen-binding Site

15
Oxygen Binding in Myoglobin

16
Preferred angles for bonding of oxygen and carbon monoxide to the iron atom of heme

19
Myoglobin

21
Mb Hb Monomer Tetromer Bind 1 O2 4 O2 Hyberbolic 141 a.a. 146 HbA (normal adult) = 22 HbF (fetal Hb) = 22 HbS (sickle cell Hb) = 2S2 HbA2 (a minor adult) = 22
 = 22. HbF (fetal Hb) = 22. HbS (sickle cell Hb) = 2S2. HbA2 (a minor adult) = 22.")
22
Hb subunits closely resemble Mb in 3D.

24
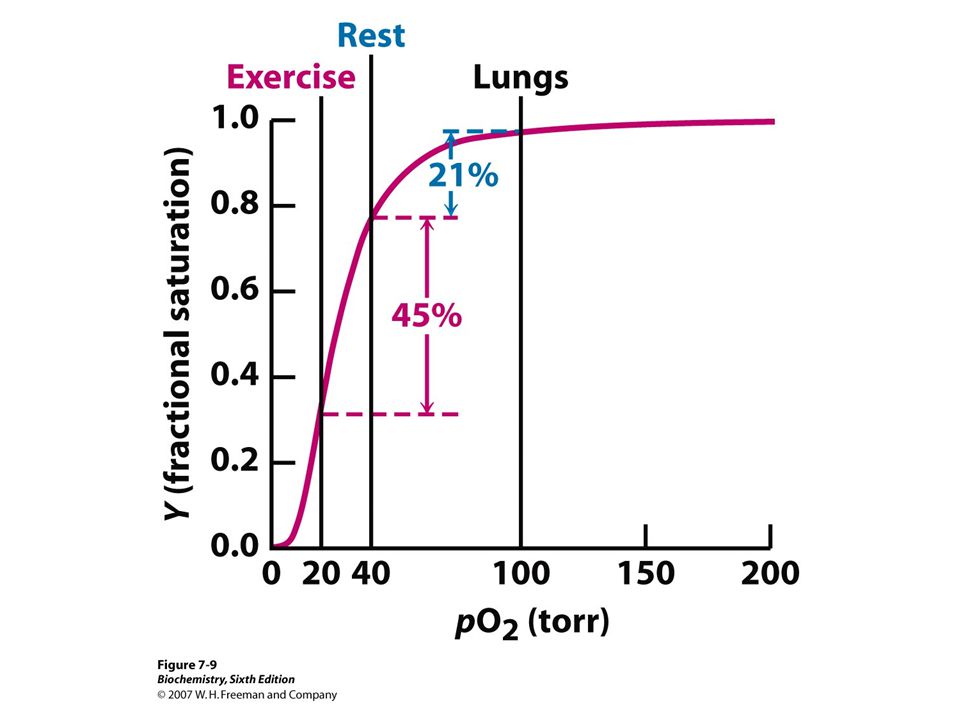
Oxygen dissociation curves of Hb and Mb.

27
Benefit of Sigmoidal Curve
Why Mb unsuitable as O2 transport protein PO2 in lung 100 mmHg PO2 n venous blood 40 mmHg It cannot serve as an effective vehicle for delivery of O2 from lungs to periphery After exercise PO2 of muscle 5 mmHg. Then Mb releases its O2 Sigmoidal curve allows Hb to deliver more O2 to tissues in response to small S in PO2

28
Salt links between and within
subunits in deoxyhemoglobin

30
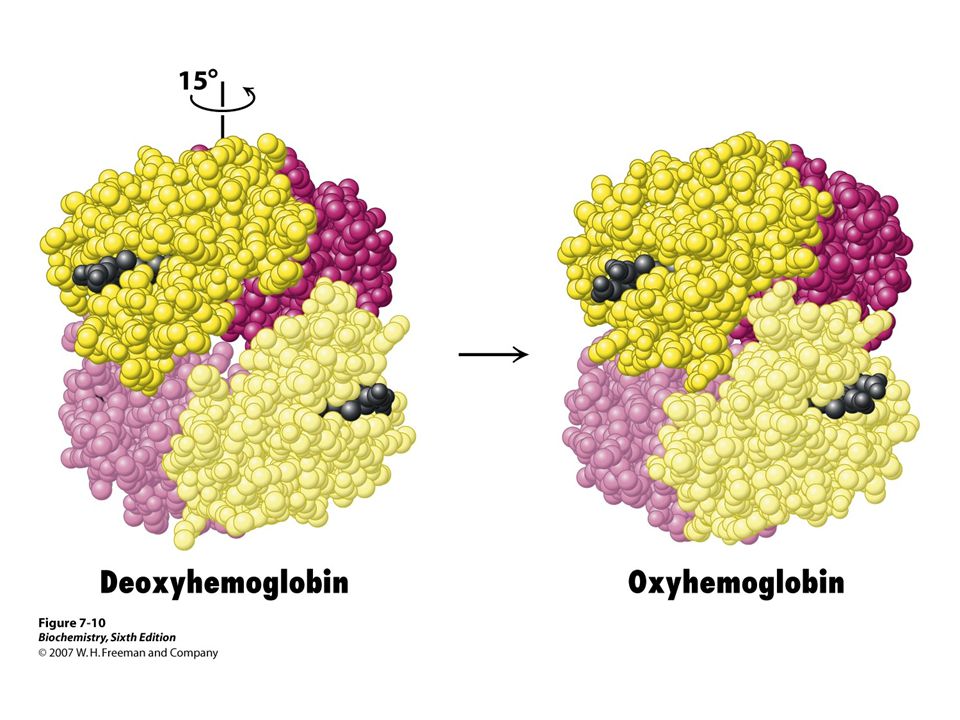
Change of quartenery structure upon oxygenation
Electron density map of oxyHb Change of quartenery structure upon oxygenation Model of oxyHb

31
Model of hemoglobin

32
Postage stamp analogy to sequential model

36
Switching of the contact region

37
Myoglobin and Hemoglobin

39
Bohr Effect

40
Chemical basis for the Bohr effect
Three amino acid residues form two Salt bridges that stabilize the T state. DeoxyHb protonation (low pH) helps this!
 helps this!")
46
Binding of BPG to deoxyHb

49
Hb has allosteric properties
Allosterism (Allos: other, Steros: space): enzyme regulation that effector binds to one site on enzyme and increases or decreases the activity of another site.
: enzyme regulation that effector binds to one site on enzyme and increases or decreases the activity of another site.")
50
Sickle Cell Anemia Genetically transmitted disease
4/1000 among blacks Sickle cell anemia homozygous Sickle cell trait is heterozygous HbS: Sickle cell Hb; HbA: Normal Hb The only difference is substitution of Val for Glu at position six of the beta chains! Fetal DNA can be analyzed for sickle cell gene Val Glu results from T A mutation Detected by new techniques Sickle cell gene has missing cleavage site for restriction endonuclease which produces 1.3 kb fragment instead 1.1 kb fragment on gel electrophoresis DNA analysis performed early in pregnancy (8 weeks gestation)
")
51
Epidemics of sickle cell anemia in Africa

54
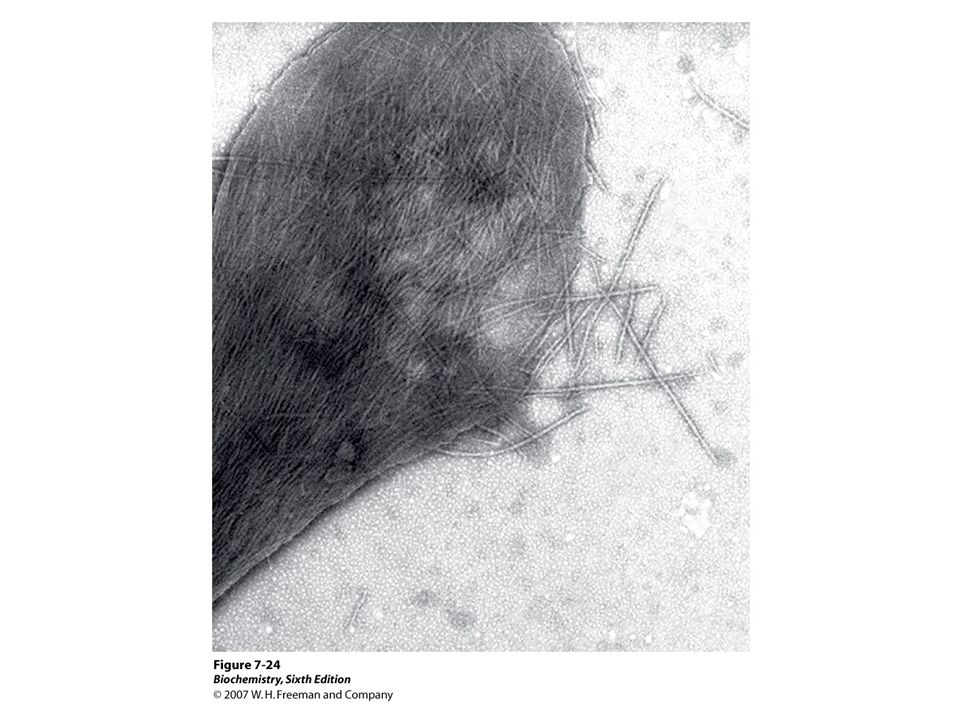
HbS

55
Why HbS aggregates? DeoxyHbS
New hydrophobic patch Val6 interacts Phe85 and Val88 of beta chain of neighboring molecule to initiate aggregation. Why aggregates do not form when HbS oxygenated In OxyHbS Phe85 and Val88 are largely buried inside Hb….

58
Ninhydrin staining is used for “fingerprinting”

59
Detect sickle cell gene by using restriction endonucleases

Similar presentations


































 Hb is found in RBCs its main function is to transport O2 to tissues. Structure: 2 parts : heme + globin Globin: four globin chains (2 α.>")


 Originally isolated from sperm whales 10X abundance greater in aquatic- than terrestrial-mammals Mb knockout.>")

