Download presentation
Presentation is loading. Please wait.

1
How will fish that evolved at constant sub-zero temperatures cope with global warming? Notothenioids as a case study Tomaso Patarnello1), Cinzia Verde2), Guido di Prisco2), Luca Bargelloni1) and Lorenzo Zane3) Bioessays 33: 260–268, 2011 WILEY Periodicals, Inc.
, Cinzia Verde2), Guido di Prisco2), Luca Bargelloni1) and Lorenzo Zane3) Bioessays 33: 260–268, 2011 WILEY Periodicals, Inc..")
2
Southern ocean

4
Interest in Antarctic organisms is, nowadays, closely related to the impact of current climate change as there is a growing concern about their ability to respond to warming. Clarke et al. recently demonstrated that the fast warming of some regions of the Southern Ocean (i.e. the Antarctic Peninsula) is already having an impact on the marine biota due to changes in sea-ice coverage. Fossil evidence indicates that many taxa have survived past climate variation and all modern species are survivors of the freezing and thawing cycles of the Pleistocene. past changes were generally much slower providing more opportunities for biological adaptation, the rates of current warming have exceeded those experienced in past millennia.
 is already having an impact on the marine biota due to changes in sea-ice coverage. Fossil evidence indicates that many taxa have survived past climate variation and all modern species are survivors of the freezing and thawing cycles of the Pleistocene. past changes were generally much slower providing more opportunities for biological adaptation, the rates of current warming have exceeded those experienced in past millennia..")
5
Most aquatic species are ectothermal, i.e. their body temperature varies with that of the environment as they lack the physiological pathways and anatomical structures needed. Antarctic fish, due to the short- and long-term stability of thermal conditions in the Southern Ocean over the last million years, are highly stenothermal. Thus, they may lack genetic variation in key adaptive traits, limiting their evolutionary potential to cope with climate changes. This work focuses on Antarctic Notothenioidei, the dominant fish sub-order. Notothenioids evolved their cold-adapted, stenothermal physiologies in the Southern Ocean over the last 20 million years, providing a unique example of marine species flock.

6
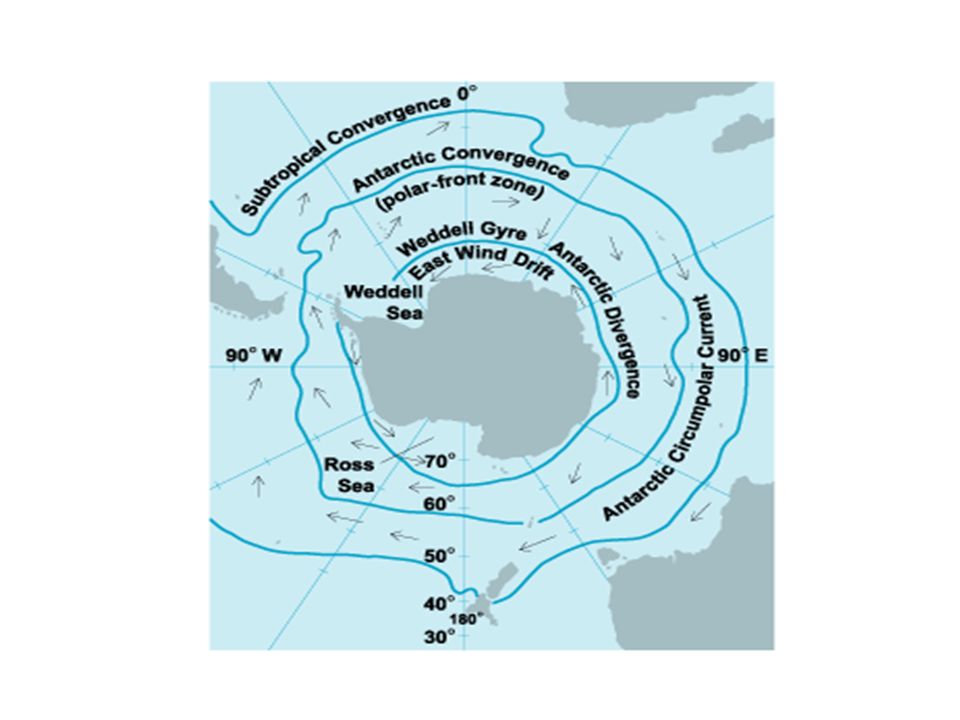
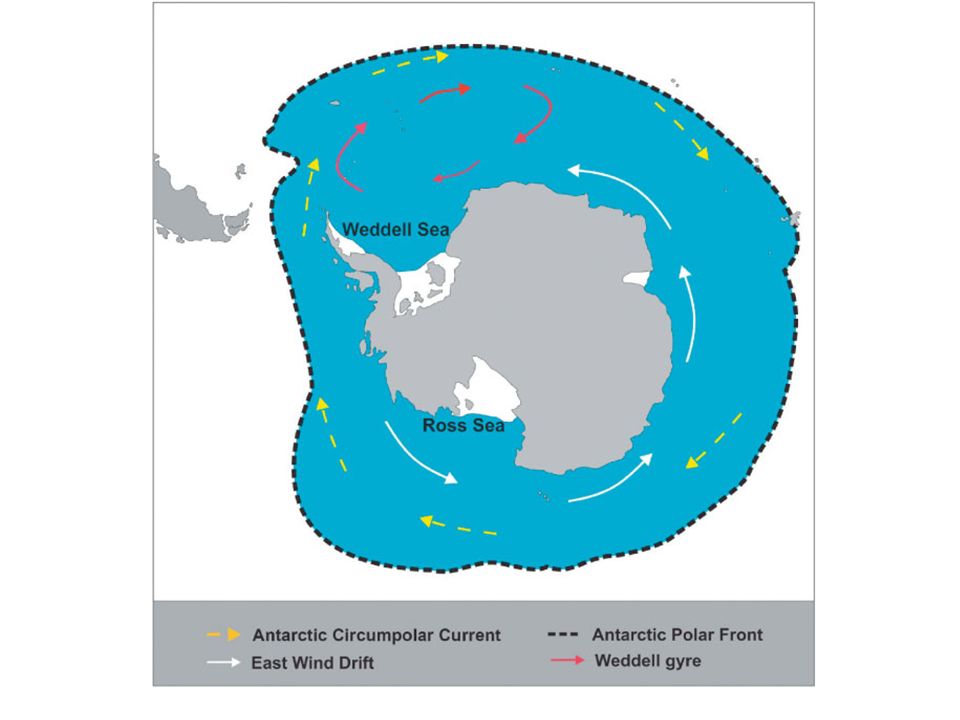
The Southern Ocean Tertiary tectonic and oceanographic events caused The complete separation of Antarctica from other continental masses The formation of the Antarctic circumpolar current (ACC) system and The gradual establishment of colder conditions.
 system and The gradual establishment of colder conditions.")
7
The ACC is a roughly circular oceanic system running between 50o and 60oS and extending to 2,000 m in depth. The northern edge is known as the Antarctic polar front (APF) and is the most intense of a series of eastward- flowing jets, representing a physical barrier to free north/south exchange of water. A sharp change in surface water temperatures (about 3 oC) marks a distinct biogeographic discontinuity.
 and is the most intense of a series of eastward- flowing jets, representing a physical barrier to free north/south exchange of water. A sharp change in surface water temperatures (about 3 oC) marks a distinct biogeographic discontinuity..")
9
The cooling of the Southern Ocean after the formation of the ACC was gradual and, the ice-sheet coverage fluctuated throughout the Cenozoic and likely produced repeated shifts of species distribution. Glacial cycles promoted northward and southward migration of the ocean fronts that define the boundaries of the Southern Ocean Antarctic planktonic ecosystems migrated towards the equator during cold periods. Thus, pelagic species could have flourished during glacial maxima due to the northward expansion of Antarctic polar waters. Benthic species, when the ice cover of the Antarctic shelf was at its maximum, bottom-living organisms could have found refugia on the shelf and slope, have migrated further into deeper waters or, have remained protected within polynyas.

10
Notothenioids: An adaptive radiation into the cold The modern Southern Ocean fish fauna consists of 322 species 88% of which are endemic. Among Antarctic fish, notothenioids underwent impressive diversification with 129 species currently assigned to this sub-order; the large majority (101) are endemic to the Southern Ocean. In high latitude shelf areas (71o–78oS), notothenioids dominate in terms of species abundance and biomass (90–95%). It is assumed that the ancestral notothenioid was benthic and that species diversification and colonisation of novel ecological niches was achieved by alteration of buoyancy, which allowed feeding in the whole water column. In the absence of a swimbladder, neutral buoyancy was obtained by a combination of reduced skeletal mineralisation and lipid deposition, leading to occupancy of pelagic, semipelagic, cryopelagic and epibenthic habitats.
 are endemic to the Southern Ocean. In high latitude shelf areas (71o–78oS), notothenioids dominate in terms of species abundance and biomass (90–95%). It is assumed that the ancestral notothenioid was benthic and that species diversification and colonisation of novel ecological niches was achieved by alteration of buoyancy, which allowed feeding in the whole water column. In the absence of a swimbladder, neutral buoyancy was obtained by a combination of reduced skeletal mineralisation and lipid deposition, leading to occupancy of pelagic, semipelagic, cryopelagic and epibenthic habitats..")
12
In addition to neutral buoyancy, notothenioids display several features considered key innovations for cold adaptation, the acquisition of antifreeze glycoproteins (AFGPs). the loss of erythrocytes and haemoglobin the whiteblooded icefish (family Channichthyidae) Cold adaptations- increased enzyme activities, modified membrane composition, lack of heat-shock response, constraints in microtubule assembly, structural and functional modifications in the oxygen transport and diffusion system, constraints in aerobic energy supply, mitochondrial functioning and anaerobic energy production
 Cold adaptations- increased enzyme activities, modified membrane composition, lack of heat-shock response, constraints in microtubule assembly, structural and functional modifications in the oxygen transport and diffusion system, constraints in aerobic energy supply, mitochondrial functioning and anaerobic energy production.")
13
Global warming: Can highly-specialised adaptations become a problem? Heat-shock response and oxidative stress transcriptional and genomic changes in response to cold adaptation. In D. mawsoni the transcriptome was compared with that of warm-water species. A set of 189 gene families were shown to be differentially expressed. (94%) were up-regulated in carp acclimated to cold, indicating the possibility of a common transcriptomic response shared by distantly related fish species. Heat-shock response, one of the most conserved biological processes across evolution, may have been lost in the adaptation to constant sub-zero temperatures. This assumption is based on evidence that Antarctic notothenioids exposed to temperatures above their natural range (+ 4 oC) do not increase production of heat-shock chaperone proteins.
 were up-regulated in carp acclimated to cold, indicating the possibility of a common transcriptomic response shared by distantly related fish species. Heat-shock response, one of the most conserved biological processes across evolution, may have been lost in the adaptation to constant sub-zero temperatures. This assumption is based on evidence that Antarctic notothenioids exposed to temperatures above their natural range (+ 4 oC) do not increase production of heat-shock chaperone proteins..")
14
The platform to probe gene expression in T. bernacchii was a cDNA microarray consisting of 9,502 full-length ESTs from multitissue libraries of the temperate eurythermal fish Gillichthys mirabilis. Despite the caution needed in drawing firm conclusions when using heterologous microarray platforms for studying expression profiling, the results confirmed the inability of T. bernacchii to up-regulate the expression of any size class of heat- shock proteins (Hsps). Hundreds of additional genes, associated with a broad range of cellular processes responsive to heat in temperate and tropical teleosts were, however, responsive in T. bernacchii. This on the one hand confirms the lack of a specific heat-shock response in Antarctic notothenioids, but on the other suggests that regulation of gene expression in response to temperature variation is only partially different between Antarctic and temperate/tropical species. while these two notothenioids have lost the ability to up-regulate hsp70 in response to short-term exposure to +4 oC, they maintain a constitutive expression of this gene under normal physiological conditions at levels equivalent to non- inducible hsc71 (a constitutive form of the 70-kDa Hsp family).
. Hundreds of additional genes, associated with a broad range of cellular processes responsive to heat in temperate and tropical teleosts were, however, responsive in T. bernacchii. This on the one hand confirms the lack of a specific heat-shock response in Antarctic notothenioids, but on the other suggests that regulation of gene expression in response to temperature variation is only partially different between Antarctic and temperate/tropical species. while these two notothenioids have lost the ability to up-regulate hsp70 in response to short-term exposure to +4 oC, they maintain a constitutive expression of this gene under normal physiological conditions at levels equivalent to non- inducible hsc71 (a constitutive form of the 70-kDa Hsp family)..")
15
An array-based comparative genomic hybridisation of Antarctic notothenioids including D. mawsoni and the phylogenetically basal (non-Antarctic) species Bovichtus variegatus and Eleginops maclovinus was recently published. Results showed that in the Antarctic species 101 protein-coding genes were duplicated from 3- to >300-fold. A large number of the overexpressed genes are involved as antioxidants, suggesting that augmented capacity for antioxidant defence is an important component of evolutionary adaptations in a cold and oxygen-rich environment. Stressful conditions such as extreme temperatures increase the production rate of reactive oxygen species (ROS) in most tissues. If this is coupled with a decreased membrane saturation typical of cold-adapted fish, it appears obvious that Antarctic organisms are more vulnerable to ROS production and oxidative damage.
 species Bovichtus variegatus and Eleginops maclovinus was recently published. Results showed that in the Antarctic species 101 protein-coding genes were duplicated from 3- to >300-fold. A large number of the overexpressed genes are involved as antioxidants, suggesting that augmented capacity for antioxidant defence is an important component of evolutionary adaptations in a cold and oxygen-rich environment. Stressful conditions such as extreme temperatures increase the production rate of reactive oxygen species (ROS) in most tissues. If this is coupled with a decreased membrane saturation typical of cold-adapted fish, it appears obvious that Antarctic organisms are more vulnerable to ROS production and oxidative damage..")
16
Loss of globin genes in the Antarctic icefish The genomic basis for haemoglobin-less condition in modern icefish resides in a large deletion of the adult ab globin locus originally identified in Chaenocephalus aceratus and Chionodraco rastrospinosus. Near, Parker and Detrich investigated the evolutionary pathway that led to gene loss, questioning whether the absence of haemoglobin expression in icefish was the result of a single mutational event prior to diversification of the extant icefish species, or it was reached via multiple mutational steps. The discovery of two distinct genomic rearrangements within the family, both leading to functional inactivation of the locus, points towards a multi-step mutational process. Previous studies had shown that the genomes of C. aceratus and C. rastrospinosus lack most of the adult ab-globin complex, retaining only a pseudogene that consists of a 3” fragment of the a-globin gene. This was assumed to be characteristic of all icefish species.

17
Similar findings were also reported for another striking feature exhibited by some icefish species, that is, loss of myoglobin (Mb) expression in oxidative muscles. Whether loss of haemoglobin and Mb was due to relaxed selection in an oxygen- rich environment or to positive selection towards the loss of oxygen-binding proteins in order to reduce blood viscosity at low temperatures, it is difficult to say. survival of T. bernacchii in spite of functional incapacitation of haemoglobin by carbon monoxide-- in the cold, stable environment of the Antarctic sea, oxygen transport is possible even in the absence of haemoglobin. Peripheral oxygen transport was evaluated in this species and compared with those of Antarctic C. aceratus and of red-blooded notothenioids inhabiting Antarctic and sub-Antarctic regions. The lowest levels of tissue oxygenation were predicted in the two icefish lacking both haemoglobin and Mb.

18
A mathematical model was applied to estimate the limits of peripheral oxygen transport imposed by anatomical (i.e. mean muscle fibre diameter, mitochondrial volume density) and physiological (mitochondrial respiration and oxygen diffusion) phenotypes of icefish as compared to red-blooded species. The purpose was to gauge the effect of any change in environmental temperature as a result of climatic change. It was shown that C. aceratus would be adversely affected by temperature perturbations, and oxidative metabolism would be impaired in C. esox during the sub-Antarctic summer maximum of 10 oC. In contrast, the model predicted that the anatomy and physiology of red-blooded notothenioids should enable them to tolerate a wide thermal range since levels of tissue oxygenation are expected to vary very little up to 20 oC. Experimental evidence supports this hypothesis. In fact, no stress response was detected in T. bernacchii upon exposure to + 3.8 oC for 4 weeks, and P. borchgrevinki was successfully acclimatised to acute and chronic (6 months) exposure to + 4 oC.
 and physiological (mitochondrial respiration and oxygen diffusion) phenotypes of icefish as compared to red-blooded species. The purpose was to gauge the effect of any change in environmental temperature as a result of climatic change. It was shown that C. aceratus would be adversely affected by temperature perturbations, and oxidative metabolism would be impaired in C. esox during the sub-Antarctic summer maximum of 10 oC. In contrast, the model predicted that the anatomy and physiology of red-blooded notothenioids should enable them to tolerate a wide thermal range since levels of tissue oxygenation are expected to vary very little up to 20 oC. Experimental evidence supports this hypothesis. In fact, no stress response was detected in T. bernacchii upon exposure to oC for 4 weeks, and P. borchgrevinki was successfully acclimatised to acute and chronic (6 months) exposure to + 4 oC..")
19
However, specimens of P. borchgrevinki infected with x-cell disease were unable to survive at + 4 oC. The latter case suggests that this species is (at least physiologically) equipped to face increasing temperature. However, concerns persist that ocean warming will nevertheless have a significant impact on its numbers because a large proportion of the natural population of this species is infected with x-cell disease. Caution is required because the responses obtained under laboratory conditions may have some important limitations in comparison to those triggered by natural conditions over a long evolutionary time.
 equipped to face increasing temperature. However, concerns persist that ocean warming will nevertheless have a significant impact on its numbers because a large proportion of the natural population of this species is infected with x-cell disease. Caution is required because the responses obtained under laboratory conditions may have some important limitations in comparison to those triggered by natural conditions over a long evolutionary time..")
20
Antifreeze proteins Unlike the uniqueness of notothenioids’ haemoglobin, antifreeze molecules have evolved in a number of organisms living at sub-zero temperatures, including invertebrates. These molecules are essential for avoiding the formation of crystalline ice in the fishes’ tissues at sub-zero temperatures, but in warmer environments they cause an unnecessary energetic cost. Decrease of serum concentration of small and large AFGPs (about 60 and 20%, respectively) was detected inP. borchgrevinki after only 4-week acclimation at + 4 oC. It should be noted that in the Arctic, where temperature is more variable, AFGPs are synthesised only during the winter. In Antarctic notothenioids, the AFGP gene evolved from a pancreatic trypsinogen- like serine-protease gene.
 was detected inP. borchgrevinki after only 4-week acclimation at + 4 oC. It should be noted that in the Arctic, where temperature is more variable, AFGPs are synthesised only during the winter. In Antarctic notothenioids, the AFGP gene evolved from a pancreatic trypsinogen- like serine-protease gene..")
21
Comparison of the gene sequences encoding AFGPs between notothenioids and the Arctic cod Boreogadus saida provided evidence of a rare example of convergent evolution. In fact, whereas the two species produce identical AFGPs, gene sequence analyses showed that the Arctic cod gene shares neither sequence identity nor gene structure with pancreatic trypsinogen; hence, it originated from an independent evolutionary pathway. Recently, Deng et al. found that type III antifreeze gene of an Antarctic zoarcid was evolved from an old sialic acid synthase following gene duplication. The evolution of this antifreeze offers another example of AFGP evolution from functionally unrelated ancestors. As the evolution of AFGPs independently occurred several times, loss of AFGP has also been observed in several notothenioids that secondarily dispersed through the APF and occupied warmer sub- or non-Antarctic waters after the major notothenioid flock was formed.

22
On the other hand, the ability of P. borchgrevinki to largely reduce AFGP serum concentration after a few weeks at + 4 oC demonstrates an effective short-term response to raised temperatures. Further experimental work is needed to determine how general this response is within the sub-order and how long the capacity to modulate AFGP expression can last.

23
Understanding notothenioids’ microevolution: From the past to the future Given the lack of fossils, insights into the speciation processes of the sub-order can be obtained only by comparing the phylogenetic relationships among species with their geographic distributions and the geological history of Antarctica. This approach indicates that speciation occurred repeatedly across both high- and sub-Antarctic biogeographic zones. during repeated glacial cycles, the habitat of benthic species may have been periodically locally eradicated due to the advance of glaciers on the shelf, restricting benthic species in refuge areas around sub-Antarctic islands or in high-productivity polynyas. It is conceivable that historical events such as periodic expansion of ice sheets in the shelf area as well as northward movement of the APF may have promoted allopatric speciation as a consequence of large fluctuations in habitat availability, periodic local extinction and fragmented refuge areas.

24
Insight into the role of climate changes, ecological factors and oceanographic barriers in shaping the genetic diversification of notothenioids can be obtained through intra-specific studies. In the pelagic notothenioid Pleuragramma antarcticum, a clear pattern of population expansion was found. This expansion was dated to about 120,000 years ago, or 200,000 years ago when incorporating the new fossil and geological evidence. Another recent intra-specific analysis detected signs of population bottlenecks and expansions possibly linked to the last glacial cycle in three species of the sub- family Trematominae. intra-specific studies provide clear evidence that notothenioids experienced drastic changes in population size during their evolutionary history. The results were suggestive of an eastward unidirectional gene flow among sampling sites consistent with the west-to-east direction of the ACC. Larval dispersal, assured by the ACC, is essential for genetic homogeneity.

25
However, in a scenario of climate change, the direction and intensity of the Southern Ocean currents might drastically change. Moreover, increased temperatures tend to reduce larval-stage duration and consequently limit the potential for dispersal. It is difficult to predict the future patterns for the Antarctic region; however, it can be anticipated that limited connectivity may ultimately lead to fragmented populations with isolated gene pools prone to genetic drift and thus exposed to the risk of polymorphism depletion. Therefore, genetic erosion of natural variation, particularly at adaptive loci, limits the evolutionary potential of Antarctic notothenioids, reducing their chances of survival in rapidly warming environments. Together, the analyses of micro-evolutionary processes indicate that Antarctic notothenioids bear the signature of historical demographic fluctuation and population fragmentation, possibly linked to climate fluctuations in the Southern Ocean over the last 100,000 years.

26
Monitoring intra-specific genetic variability of Southern Ocean marine species is important because climate change in this extremely delicate and vulnerable area of the planet may result in loss of genetic variation for several reasons. First, climate change can reduce the available habitats for species or push them into sub-optimal conditions at the margins of their physiological capacity to perform essential functions, thus producing a significant reduction in population size or even local extinction with potential impact on species polymorphism. Second, an increase of temperatures may change oceanographic circulation and may alter the extent and time of release of larvae into the current system, ultimately disrupting the complex mechanisms of recruitment and connectivity between locations. Third, climate change has been shown to have significant effects on species distributions.

27
Whether or not a scenario of forced sympatric distributions may cause competitive exclusion and inter-specific hybridisation is not easily predictable, but it certainly points to the need for future research. Finally, climate change exposes the Southern Ocean to invasions of non indigenous taxa that may act as competitors to indigenous species, destabilising ecosystems and causing community shifts, as envisaged for benthos invertebrates. This scenario raises important concerns given the peculiarities of Antarctic organisms (e.g. long generation time and low fecundity), which make them weak competitors. Human harvesting of several Antarctic fish species further complicates this problem.
, which make them weak competitors. Human harvesting of several Antarctic fish species further complicates this problem..")
Similar presentations



 c and z are fitting parameters.>")







, ecology (e.g., local.>")


 The McGraw-Hill Companies, Inc.>")





