Download presentation
Presentation is loading. Please wait.

1
The Nucleus 9

2
9 The Nucleus Chapter Outline The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Internal Organization of the Nucleus The Nucleolus and rRNA Processing

3
Introduction The nucleus is the principal feature that distinguishes eukaryotic from prokaryotic cells. It serves as the repository of genetic information and as the cell’s control center. Separation of the genome from the site of mRNA translation thus plays a central role in eukaryotic gene expression.

4
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm The nuclear envelope separates the contents of the nucleus from the cytoplasm. The selective traffic of proteins and RNAs through nuclear pore complexes establishes the nuclear composition and plays a critical role in regulating eukaryotic gene expression.

5
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm The nuclear envelope consists of two membranes, an underlying nuclear lamina, and nuclear pore complexes. The outer membrane is continuous with the endoplasmic reticulum. It is enriched in membrane proteins that bind the cytoskeleton. The inner membrane has proteins that bind the nuclear lamina.

6
Figure 9.1 The nuclear envelope (Part 1)
")
7
Figure 9.1 The nuclear envelope (Part 2)
")
8
Figure 9.1 The nuclear envelope (Part 3)
")
9
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Each nuclear membrane is a phospholipid bilayer permeable only to small nonpolar molecules. Nuclear pore complexes are the sole channels for small polar molecules, ions, proteins, and RNA to pass through the nuclear envelope.

10
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm The nuclear lamina is a fibrous mesh that provides structural support. It consists of fibrous proteins called lamins, and other proteins. Mutations in lamin genes cause several different inherited tissue-specific diseases.

11
Figure 9.3 Electron micrograph of the nuclear lamina

12
Molecular Medicine 9.1 Nuclear Lamina Diseases: (A) A child with Hutchinson-Gilford progeria. (B) Intron-exon structure of the LMNA gene and lamin A protein
 Intron-exon structure of the LMNA gene and lamin A protein.")
13
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Mammalian cells have three lamin genes, (A, B, and C), which code for at least seven proteins. Two lamins interact to form a dimer in which the α-helical regions of two polypeptide chains wind around each other to form a coiled coil. The lamin dimers associate with each other to form the nuclear lamina.

14
Figure 9.4 Model of lamin assembly

15
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Lamins bind to inner nuclear membrane proteins such as emerin and lamin B receptor. The lamins and lamin-associated proteins also bind to chromatin. Lamins also extend in a loose meshwork throughout the interior of the nucleus.

16
Figure 9.5 The nuclear lamina

17
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Nuclear pore complexes are large, complex structures. In vertebrates, they is composed of about 30 different pore proteins (nucleoporins). The control of molecular traffic between the nucleus and the cytoplasm plays a fundamental role in the physiology of all eukaryotic cells.
. The control of molecular traffic between the nucleus and the cytoplasm plays a fundamental role in the physiology of all eukaryotic cells..")
18
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Molecules travel through pore complexes by two mechanisms: Passive transport—small molecules pass freely in either direction through open aqueous channels. Macromolecules (proteins and RNAs) are transported by a selective, energy-dependent mechanism.
 are transported by a selective, energy-dependent mechanism..")
19
Figure 9.6 Molecular traffic through nuclear pore complexes

20
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Electron microscopy shows that pore complexes have eightfold symmetry organized around a large central channel.

21
Figure 9.7 Electron micrograph of nuclear pore complexes

22
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm A nuclear pore complex consists of eight spokes connected to rings at the nuclear and cytoplasmic surfaces. The spoke-ring assembly surrounds a central channel. Protein filaments extend from the rings, forming a basketlike structure on the nuclear side.

23
Figure 9.8 Model of the nuclear pore complex

24
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Proteins that must enter the nucleus have amino acid sequences called nuclear localization signals. These are recognized by nuclear transport receptors which direct transport of the proteins through the nuclear pore complex.

25
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Nuclear localization signals were first identified in 1984, using a viral replication protein SV40 T antigen. Using T antigen mutants, they determined the amino acid sequence that was responsible for nuclear localization. When the same sequence was attached to other proteins, they also were transported to the nucleus.

26
Key Experiment 9.1 Identification of Nuclear Localization Signals: Determining cellular localization of fusion proteins

27
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm The T antigen nuclear localization signal is a single stretch of amino acids. Others are bipartite, consisting of two amino acids sequences separated by another amino acid sequence.

28
Figure 9.9 Nuclear localization signals

29
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Nuclear localization signals are recognized by nuclear transport receptors called importins. Activity of nuclear transport receptors is regulated by Ran, a GTP-binding protein.

30
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm For Ran, enzymes for GTP hydrolysis to GDP are on the cytoplasmic side of the nuclear envelope; enzymes for exchange of GDP for GTP are on the nuclear side. This leads to higher concentration of Ran/GTP in the nucleus, and determines the directionality of transport.

31
Figure 9.10 Distribution of Ran/GTP across the nuclear envelope

32
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm A specific importin binds to the nuclear localization signal of a cargo protein in the cytoplasm. This complex binds to the cytoplasmic filaments of the pore complex. Transport proceeds by sequential binding to specific nuclear pore proteins located further and further toward the nuclear side of the pore complex.

33
Figure 9.11 Protein import through the nuclear pore complex

34
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Nucleoporins with multiple Phe-Gly repeats (FG-proteins) line the central channel. At the nuclear side the cargo/importin complex is disrupted by binding of Ran/GTP. This causes a conformation change in the importin, which releases the cargo protein into the nucleus.

35
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm The importin-Ran/GTP complex is then exported back to the cytoplasm where the GTP is hydrolyzed to GDP. The importin is released and can participate in another round of transport. Ran/GDP is transported back to the nucleus by its own import receptor (NTF2), where Ran/GTP is regenerated.
, where Ran/GTP is regenerated..")
36
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Proteins are targeted for export from the nucleus by specific amino acid sequences, called nuclear export signals. These signals are recognized by receptors in the nucleus (exportins), which direct protein transport to the cytoplasm.
, which direct protein transport to the cytoplasm..")
37
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Many importins and exportins are members of a family of nuclear transport receptors known as karyopherins.

39
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Ran is required for nuclear export as well as import. Ran/GTP promotes binding of exportins and their cargo proteins, but dissociates complexes between importins and their cargos.

40
Figure 9.12 Nuclear export

41
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Protein transport is another point at which nuclear protein activity can be controlled; for example regulation of import and export of transcription factors. Changes in receptor affinity of only two pore complex proteins contributed to the evolutionary split between Drosophila melanogaster and Drosophila simulans.

42
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm In one mechanism, transcription factors (or other proteins) associate with cytoplasmic proteins that mask their nuclear localization signals, and so they remain in the cytoplasm.
 associate with cytoplasmic proteins that mask their nuclear localization signals, and so they remain in the cytoplasm.")
43
Figure 9.13 Regulation of nuclear import of transcription factors

44
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Other transcription factors are regulated by phosphorylation. Example: the yeast transcription factor Pho4 is phosphorylated at a site adjacent to its nuclear localization signal, which interferers with its import.

45
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Most RNAs are exported from the nucleus to the cytoplasm for use in protein synthesis. It is an active, energy-dependent process requiring the transport receptors to interact with the nuclear pore complex.

46
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm RNAs are transported as ribonucleoprotein complexes (RNPs). rRNAs are associated with ribosomal proteins and specific RNA processing proteins in the nucleolus. mRNAs are associated with at least 20 proteins during processing and eventual transport to the cytoplasm.

47
Figure 9.14 Transport of a ribonucleoprotein complex

48
The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm Many small RNAs (snRNAs and snoRNAs) function within the nucleus. snRNAs are transported to the cytoplasm by an exportin (Crm1), where they associate with proteins to form snRNPs and return to the nucleus.
, where they associate with proteins to form snRNPs and return to the nucleus..")
49
Figure 9.15 Transport of snRNAs between nucleus and cytoplasm

50
Internal Organization of the Nucleus The nucleus has an internal structure that organizes the genetic material and localizes nuclear functions. In animal cells, lamins serve as sites of chromatin attachment and organize other proteins into functional nuclear bodies.

51
Internal Organization of the Nucleus During interphase, heterochromatin remains highly condensed and is transcriptionally inactive; the euchromatin is decondensed and distributed throughout the nucleus.

52
Figure 9.16 Heterochromatin in interphase nuclei

53
Internal Organization of the Nucleus Two types of heterochromatin: Constitutive heterochromatin contains DNA that is not transcribed, such as satellite sequences at centromeres. Facultative heterochromatin contains sequences that are not transcribed in the cell being examined but are transcribed in other cell types.

54
Internal Organization of the Nucleus During interphase, chromosomes are organized and divided into discrete functional domains. This organization was first suggested in 1885 and confirmed in 1984 by studies of polytene chromosomes in Drosophila salivary glands.

55
Figure 9.17 Chromosome organization

56
Internal Organization of the Nucleus Each chromosome occupies a discrete region of the nucleus. They are closely associated with the nuclear envelope at many sites. Many of these associations result in repression of gene expression but some promote transcription of specific genes.

57
Figure 9.18 Organization of Drosophila chromosomes

58
Internal Organization of the Nucleus In mammalian cell nuclei, actively transcribed genes are localized to the periphery of the territories. RNA processing and transport are thought to occur in channels (interchromosomal domains) that separate the chromosomes.
 that separate the chromosomes..")
59
Figure 9.19 Organization of chromosomes in the mammalian nucleus

60
Internal Organization of the Nucleus Different cell types express different genes, so their facultative heterochromatin is different, and varying regions of the chromosomes interact with the nuclear lamina in different cells and tissues.

61
The Hilbert curve is a one-dimensional fractal trajectory that densely fills higher-dimensional space without crossing itself. Leiberman-Aiden et al. (2009) Science 326:289-293
 Science 326:")
62
3-D structure of human genome: fractal globule architecture

63
Internal Organization of the Nucleus In interphase, the chromatin is organized into looped domains containing about 50–100 kb of DNA. In amphibian oocytes, actively transcribed regions of DNA can be seen as large loops of extended chromatin.

64
Figure 9.20 Looped chromatin domains

65
Internal Organization of the Nucleus Most nuclear processes are localized to distinct regions of the nucleus. Many proteins are localized in low- density, sponge-like subnuclear bodies that allow macromolecules to move in and out. The nature and function of these nuclear substructures are not yet clear.

66
Internal Organization of the Nucleus Mammalian cell nuclei contain clustered sites of DNA replication. They are identified by labeling newly synthesized DNA with bromodeoxyuridine, which is incorporated in place of thymidine. Immunofluorescence shows the newly replicated DNA is present in discrete clusters.

67
Figure 9.21 Clustered sites of DNA replication

68
Internal Organization of the Nucleus DNA replication appears to take place in distinct functional bodies with multiple replication complexes. These have been called replication factories.

69
Internal Organization of the Nucleus The mRNA splicing machinery is concentrated in discrete nuclear bodies, called nuclear speckles. They can be seen with immunofluorescent staining with antibodies against snRNPs and splicing factors.

70
Figure 9.22 Localization of splicing components

71
Internal Organization of the Nucleus PML bodies were first identified as localization sites of a transcriptional regulatory protein involved in acute promyelocytic leukemia (PML). They are sites of accumulation of transcription factors and chromatin- modifying proteins, but their function remains unknown.

72
Figure 9.23 A PML body

73
Internal Organization of the Nucleus Cajal bodies contain the characteristic protein coilin and are enriched in small RNPs. They may function as sites of RNP assembly and processing.

74
Figure 9.24 Cajal bodies in the nucleus

75
The Nucleolus and rRNA Processing The nucleolus is the site of rRNA transcription and processing, and some aspects of ribosome assembly. Cells need large numbers of ribosomes at specific times for protein synthesis. Actively growing mammalian cells have 5 to 10 million ribosomes that must be synthesized each time the cell divides.

76
The Nucleolus and rRNA Processing The nucleolus is not surrounded by a membrane. The 5.8S, 18S, and 28S rRNAs are transcribed as a single unit in the nucleolus by RNA polymerase I, yielding a 45S ribosomal precursor RNA.

77
Figure 9.25 Ribosomal RNA genes

78
The Nucleolus and rRNA Processing All cells contain multiple copies of the rRNA genes. In oocytes, the rRNA genes are amplified to support synthesis of ribosomes needed for early development. rRNA genes are amplified two-thousand- fold in Xenopus oocytes, in thousands of nucleoli, resulting in 10 12 ribosomes per oocyte.

79
Figure 9.26 Nucleoli in amphibian oocytes

80
The Nucleolus and rRNA Processing Nucleoli have three regions: the fibrillar center, dense fibrillar component, and granular component. They are thought to represent sites of progressive stages of rRNA transcription, processing, and ribosome assembly.

81
Figure 9.27 Structure of the nucleolus

82
The Nucleolus and rRNA Processing Following each cell division, nucleoli become associated with the nucleolar organizing regions that contain the 5.8S, 18S, and 28S rRNA genes. Transcription of 45S pre-rRNA leads to fusion of small prenucleolar bodies. In most cells, the initially separate nucleoli then fuse to form a single nucleolus.

83
The Nucleolus and rRNA Processing Each nucleolar organizing region contains a cluster of tandemly repeated rRNA genes separated by nontranscribed spacer DNA. The genes and their growing RNA chains can be seen in electron micrographs.

84
Figure 9.28 Transcription of rRNA genes

85
The Nucleolus and rRNA Processing In higher eukaryotes the primary transcript of rRNA genes is the large 45S pre-rRNA. The pre-rRNA is processed via a series of cleavages, which is similar in all eukaryotes.

86
Figure 9.29 Processing of pre-rRNA

87
The Nucleolus and rRNA Processing Processing of pre-rRNA also includes addition of methyl groups to bases and ribose residues, and conversion of uridine to pseudouridine.

88
The Nucleolus and rRNA Processing Nucleoli contain over 300 proteins and many small nucleolar RNAs (snoRNAs) that function in pre-rRNA processing. snoRNAs are complexed with proteins, forming snoRNPs. They assemble on pre-rRNA to form processing complexes similar to the formation of spliceosomes on pre-mRNA.

89
The Nucleolus and rRNA Processing Most snoRNAs guide RNAs to direct specific base modifications of pre-rRNA. They contain short sequences of nucleotides that are complementary to 18S or 28S rRNA and include the sites of base modification in the rRNA.

90
Figure 9.30 Role of snoRNAs in base modification of pre-rRNA

91
The Nucleolus and rRNA Processing Formation of ribosomes requires assembly of pre-rRNA with ribosomal proteins and 5S rRNA. Ribosomal proteins are imported to the nucleolus from the cytoplasm, where they assemble with the pre-rRNA prior to cleavage.

92
The Nucleolus and rRNA Processing 5S rRNAs are similarly assembled into preribosomal particles elsewhere in the nucleolus. Additional ribosomal proteins and the 5S rRNA are incorporated as cleavage and processing proceeds. The two nascent ribosomal subunits separate and are exported to the cytoplasm.

93
Figure 9.31 Ribosome assembly

94
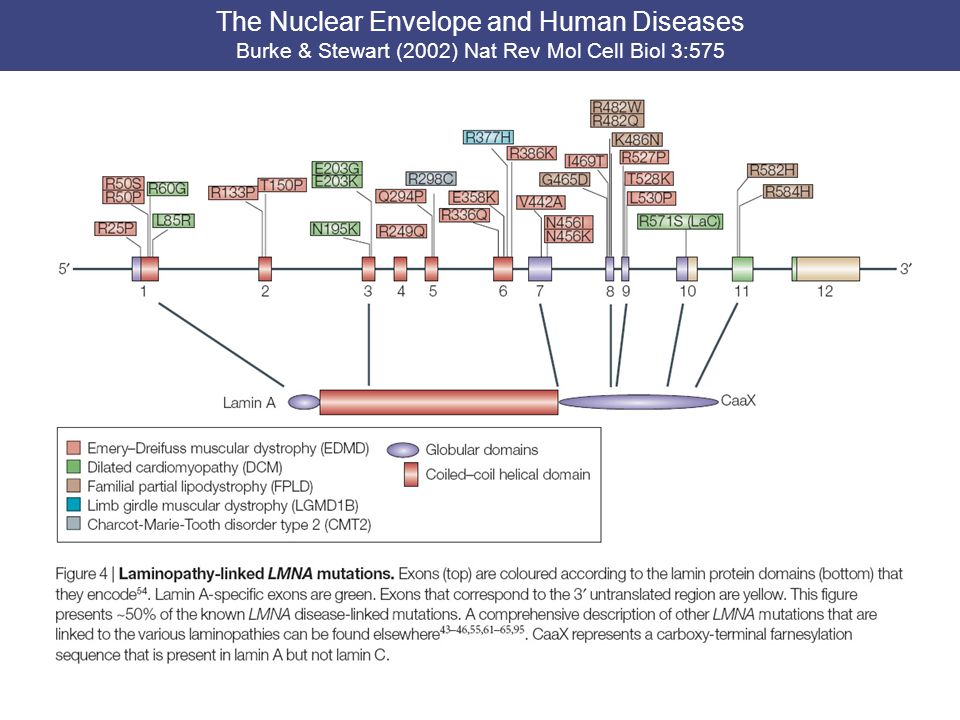
The Nuclear Envelope and Human Diseases Burke & Stewart (2002) Nat Rev Mol Cell Biol 3:575 NE is essential in defining higher-order nuclear structure by providing anchoring sites for chromatin domains at the nuclear periphery. “Laminopathies” include cardiac and skeletal myopathies, partial lipodystrophy (loss of adipocytes), peripheral neuropathy. NE not only has a ubiquitous role in the maintenance of nuclear architecture, but also might provide cell-specific functions.
, peripheral neuropathy. NE not only has a ubiquitous role in the maintenance of nuclear architecture, but also might provide cell-specific functions..")
95
The Nuclear Envelope and Human Diseases Burke & Stewart (2002) Nat Rev Mol Cell Biol 3:575
 Nat Rev Mol Cell Biol 3:575")
97
Laminopathies: defects in genes that encode nuclear-lamina and lamina-associated proteins – specifically A-type lamins and emerins A mutation in the emerin gene leading to a loss of emerin from the nuclear periphery Childhood onset of progressive muscle wasting and weakening Early contractures of the Achilles tendons, tendons of the elbows and necks…. Late-onset symptoms: abnormal cardiac rhythms, conduction block, and cardiomyopathy… X-linked Emery-Dreifuss muscular dystrophy

98
Mutations in Lamin A gene (LMNA) mostly missense mutations affecting both Lamin A and Lamin C proteins “Autosomal-dominant” means that it is haploinsufficient. Mouse model with Lmna deletion: a human EDMD model Autosomal-dominant muscular dystrophy The Nuclear Envelope and Human Diseases Burke & Stewart (2002) Nat Rev Mol Cell Biol 3:575
 Nat Rev Mol Cell Biol 3:575.")
99
Lmna knockout mouse model The Nuclear Envelope and Human Diseases Burke & Stewart (2002) Nat Rev Mol Cell Biol 3:575
 Nat Rev Mol Cell Biol 3:575")
100
Lmna knockout mouse model The Nuclear Envelope and Human Diseases Burke & Stewart (2002) Nat Rev Mol Cell Biol 3:575 Anti-LaminB Anti-LaminA Anti-Emerin LaminA 를 발현시켜 주니 Emerin 이 제대로 localize 된다.
 Nat Rev Mol Cell Biol 3:575 Anti-LaminB Anti-LaminA Anti-Emerin LaminA 를 발현시켜 주니 Emerin 이 제대로 localize 된다.")
101
Mutations in Lamin A gene (LMNA) One mutation is a frameshift at codon 320. This mutation is predicted to produce lamin proteins without NLS and are defective in assembly. Skeletal myopathy and cardiac conduction defects Skeletal and cardiomyopathy The Nuclear Envelope and Human Diseases Burke & Stewart (2002) Nat Rev Mol Cell Biol 3:575
 Nat Rev Mol Cell Biol 3:575.")
102
Mutations in Lamin A gene (LMNA) Charcot-Marie-Tooth type 2 (CMT2) syndrome R298C This mutation is likely to perturb the lateral interactions between A- type lamins. Symptoms: muscle weakness accompanied by loss of large myelinated muscle fibers and axonal degeneration Neuropathy The Nuclear Envelope and Human Diseases Burke & Stewart (2002) Nat Rev Mol Cell Biol 3:575
 Nat Rev Mol Cell Biol 3:575.")
103
Mutations in Lamin A gene (LMNA), mainly in exon 11 Dunnigan-type familial partial lipodystrophy (FPLD) Changes in fat distribution (more in upper body, face, and neck) High TG, high insulin, insulin-resistant diabetes Lmna mutant mice do not show such phenotype. Thus, FPLD is not caused simply by loss of some lamin function. R482W, maybe a gain-of-function mutation? Lipodystrophy The Nuclear Envelope and Human Diseases Burke & Stewart (2002) Nat Rev Mol Cell Biol 3:575
 Nat Rev Mol Cell Biol 3:575.")
105
Nuclear fragility Weak nuclear envelope leading to nuclear damage and cell death Muscle contraction – mechanical stress on weak NE Lipodystrophy 는 mechanical stress 와 무관 Disease Mechanisms The Nuclear Envelope and Human Diseases Burke & Stewart (2002) Nat Rev Mol Cell Biol 3:575 Gene-expression effects Heterochromatin 의 구성을 바꾸어 gene expression pattern 에 변화가 온다 ?
 Nat Rev Mol Cell Biol 3:575 Gene-expression effects Heterochromatin 의 구성을 바꾸어 gene expression pattern 에 변화가 온다")
106
Premature aging LMNA mutation (Nature, 2003) Processing of Prelamin A to Lamin A is affected. Farnesylated prelamin A accumulates in the nuclear envelope. Farnesyltransferase inhibitor (FTI) to release prelamin A from NE partially reverses the nuclear damage. Phase II clinical trial ongoing Hutchinson-Gilford Progeria Syndrome treated Untreated Nuclear LMNA blebbing
 to release prelamin A from NE partially reverses the nuclear damage. Phase II clinical trial ongoing Hutchinson-Gilford Progeria Syndrome treated Untreated Nuclear LMNA blebbing.")
Similar presentations





















 1. Nuclear envelope 2.Nucleoskeleton 3.Nuclear.>")