Download presentation
Presentation is loading. Please wait.

2
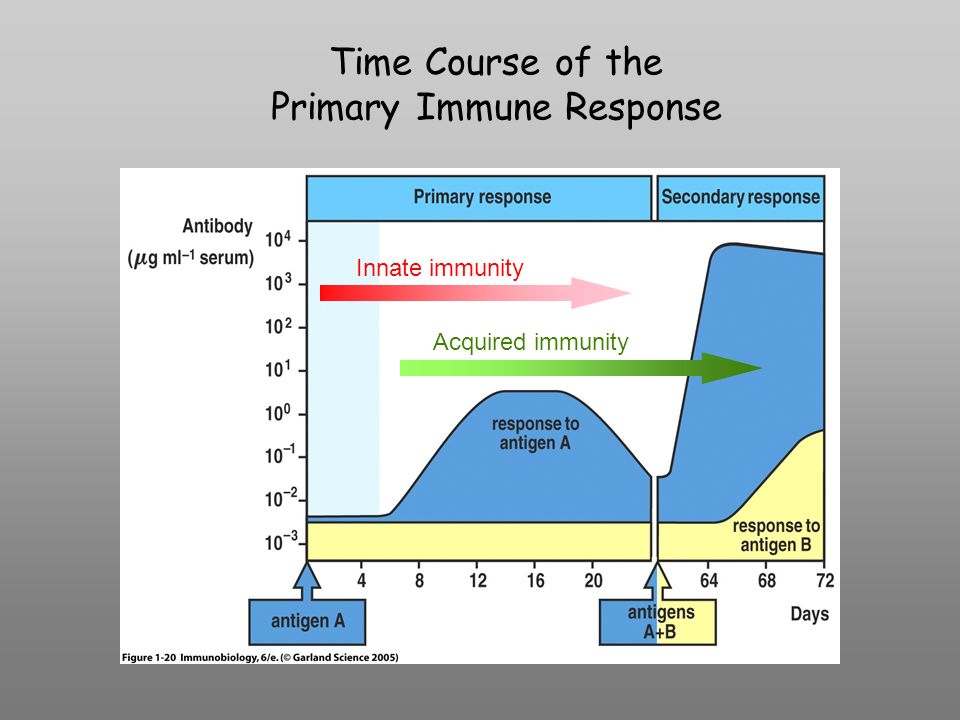
Time Course of the Primary Immune Response Innate immunity Acquired immunity

3
Ig Maturation

4
Journey of a B Cell

5
Contact Between the TCR and MHC/peptide: Not All Peptides are Created Equal

6
The Two-Signal Theory of T-cell Activation APC = Antigen-presenting cells TCR = T-cell receptor for antigen DC = Dendritic cell CD80 = Co-stimulatory receptor 2 1 12 No response or Anergy Activation

7
V C C V peptide CD3 TCR CD4 MHC II V C C V CD3 TCR MHC I CD8 (1) Interacts with MHC class II expressing cells (APCs) (2) Helps B cells to synthesize antibody (3) Induces and activates macrophages (4) Secretes cytokines (1) Interacts with MHC class I-expressing cells (all nucleated cells) (2) Kill MHC class I-expressing target cells (3) Secretes cytokines CD4+ T cell CD8+ T cell peptide Two Major Functional T Cell Subsets Lck APC
 Interacts with MHC class II expressing cells (APCs) (2) Helps B cells to synthesize antibody (3) Induces and activates macrophages (4) Secretes cytokines (1) Interacts with MHC class I-expressing cells (all nucleated cells) (2) Kill MHC class I-expressing target cells (3) Secretes cytokines CD4+ T cell CD8+ T cell peptide Two Major Functional T Cell Subsets Lck APC")
8
CD4+ T Cells Activate Macrophages and B cells

9
Immunity Tolerance Activation Suppression Regulation of the Immune Response: a Conceptual View

10
Immunity Tolerance Activation Suppression Regulation of the Immune Response: a Conceptual View Autoimmunity Immunodeficiency

11
Immunity Tolerance Activation Suppression Regulation of the Immune Response: a Conceptual View Autoimmunity Immunodeficiency

12
Antibodies: Secreted or Transmembrane TCR: Transmembrane

13
Figure 4-2 DNA Rearrangement RemovesSequences Between V, D and J Segments RNA Splicing Removes Sequences Between J and C Segments DNA RNA

15
Antigen-Independent B-Cell Development 1.DNA rearrangements establish the primary repertoire, creating diversity 2. Allelic exclusion ensures that each clone expresses a single antibody on the surface, establishing specificity 3. Deletion of self-reactive clones establishes tolerance Bone Marrow

16
THE B CELL RECEPTOR 1.Bound antigen is inter- nalized and presented to T cells. 2. Bound antigen gives signals to the B cell to proliferate and differentiate.

17
Signalling from the BCR Lack of Btk causes Bruton’s X-linked agammaglobulinemia (blocked at pre-B stage)
")
18
Antigen-Dependent B Cell Development In Periphery (spleen and LN) Antigen and T H cells give B cells two signals: 1) proliferate 2) differentiate T-cell dependent responses are refined two ways: 1) higher affinity antibodies 2) IgG/A/E (“switched”) isotypes Two products of B cell development: 1) plasma cells secrete Ig (final effector) 2) memory cells respond to II o antigen
 Antigen and T H cells give B cells two signals: 1) proliferate 2) differentiate T-cell dependent responses are refined two ways: 1) higher affinity antibodies 2) IgG/A/E ( switched ) isotypes Two products of B cell development: 1) plasma cells secrete Ig (final effector) 2) memory cells respond to II o antigen")
20
T Cell-B Cell Communication 1.Cell-cell signals from CD40L/CD40 and other surface molecules. 2. Secreted cytokines (B cells signal T cells by presenting Ag in association with MHC II) T cells provide 2 kinds of help to B cells:
 T cells provide 2 kinds of help to B cells:.")
21
The Germinal Center 1.Affinity maturation a. Somatic hypermutation b. Selection for high affinity clones 2. Isotype switch recombination 4. Final maturation to memory or plasma cell. 3. Peripheral tolerance

22
Proliferation Ag(FDC) + T cell help (Iterative cycles) SURVIVAL but T help and no Ag (eliminates low affinity clones) or Ag and no T help (eliminates self-reactive clones, giving tolerance) DEATH + Somatic Hypermutation AFFINITY MATURATION IN THE GC Dark zone Light zone
 + T cell help (Iterative cycles) SURVIVAL but T help and no Ag (eliminates low affinity clones) or Ag and no T help (eliminates self-reactive clones, giving tolerance) DEATH + Somatic Hypermutation AFFINITY MATURATION IN THE GC Dark zone Light zone")
23
1. Memory B cells Surface Ig, usually IgG High affinity for antigen Long-lived, even in the absence of antigen Respond rapidly to secondary stimulation 2. Plasma Cells Secrete copious amounts of Ig, no surface Ig Non-dividing Some are short-lived, some become long-lived in the bone marrow

24
Secreted Antibodies Function in Various Ways To Eliminate Foreign Invaders

25
Notion of immunological “SELF” Skin graft transplantation compatibility Graft compatibility genetically determined Extremely polymorphic trait -many alleles Governed by genes of major histocompatibility complex (MHC) that encode MHC molecules, the principal targets of rejection Rejection by adaptive immune system T cell response Differences in MHC molecules between individuals are central to determining “SELF” How T cells recognize antigen
 that encode MHC molecules, the principal targets of rejection Rejection by adaptive immune system T cell response Differences in MHC molecules between individuals are central to determining SELF How T cells recognize antigen")
26
The selection process (Thymic “education”) has two stages First stage selects clones capable of recognizing self peptide in an individual’s own MHC molecules - positive selection Second stage eliminates overtly self reactive clones with high affinity for self peptide:MHC- negative selection (Self-peptides are used as a surrogate for foreign peptides since there are few non-self peptides) ***
 has two stages First stage selects clones capable of recognizing self peptide in an individual’s own MHC molecules - positive selection Second stage eliminates overtly self reactive clones with high affinity for self peptide:MHC- negative selection (Self-peptides are used as a surrogate for foreign peptides since there are few non-self peptides) ***")
27
The specificity of self/non-self peptide binding to MHC molecules determined by pockets that only bind certain amino acid side chains MHC genes are extremely polymorphic and alleles encode pockets with specificities for different amino acid side chains The TCR is specific for both peptide and MHC- A complex ligand Polymorphic residues of MHC The TCR repertoire differs from individual to individual The definition of immunologic self is made by selecting the clonal T cell repertoire on self-peptides bound to the individual’s particular allelic forms of MHC molecules

28
Cytosolic Virus or Pathogen Ingested Bacteria or Endocytic Pathogen Extracellular Pathogen or Toxin Peptide degraded in: Peptides bind to: Presented to: Effect on presenting cell of T cell recognition: Cytosol MHC class I CD8 T cells Death of cell presenting the viral antigen Endocytic vesicles MHC class II CD4 T cells Provision of help to B cell for production of antibodies Endocytic vesicles MHC class II (or I) CD4 T cells (or CD8) Activation of cell to enhance pathogen killing Challenge: Any cellMacrophage/DCB cell The immune system makes this distinction by loading and recognizing peptides in either class I or class II MHC Presenting cell:
 CD4 T cells (or CD8) Activation of cell to enhance pathogen killing Challenge: Any cellMacrophage/DCB cell The immune system makes this distinction by loading and recognizing peptides in either class I or class II MHC Presenting cell:")
29
Structure of peptide-binding class I MHC domain N 11 22

31
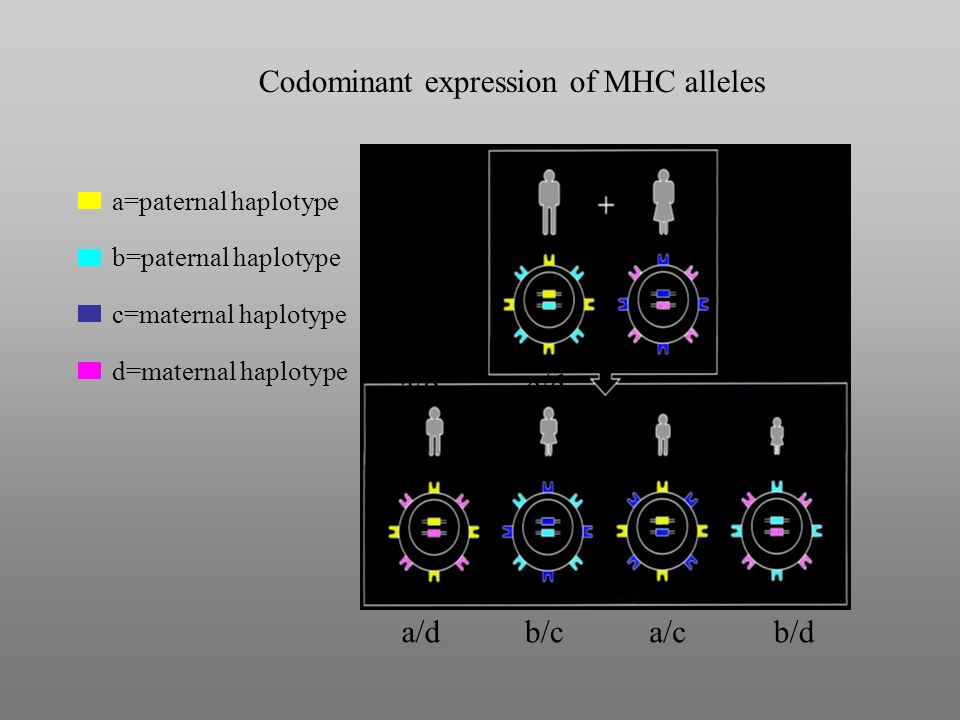
Codominant expression of MHC alleles a/b c/d a/db/ca/cb/d a=paternal haplotype b=paternal haplotype c=maternal haplotype d=maternal haplotype

32
Polymorphic amino acids that distinguish alleles of MHC class I molecules are found primarily in pockets that determine peptide binding or on the surface that interacts with the TCR

33
Because the TCR recognizes both peptide and MHC molecule, T cell recognition of MHC-peptide is both MHC restricted and specific for the immunizing peptide Peptide from virus X T cell APC HLA- B7 Peptide from virus X APC HLA- B27 T cell Peptide from virus Y T cell APC HLA- B7 Target killed: YesNo In each of the 3 experiments the T cell is from a HLA-B7 person who recovered from infection by virus “X”. The APC target cell is either infected with virus X or Y and is from an individual who is either HLA-B7 or HLA-B27

34
CD4 Staining CD8 Staining CD4- CD8- CD4+CD8 - CD4+CD8 + CD4-CD8+ 3% 5%80% 12% TCR repertoire selection and thymocyte differentiation into CD4+ or CD8+ T cells

35
Individuals with different MHCs have different TCR repertoires T cells mature into CD4 or CD8 single-positive cells as a result of positive selection. Implications of Positive/Negative Selection

36
MHC class II/ autopeptide TCR CD3 CD80 CD40 MHC class II CD40 CD40L CD28CD80 Activated T cell (1) induction of cytokines/chemokines (IL-8, IL-12, TNF- , MIP-1 ) (2) stimulation of CD80 and CD86 expression and co- stimulatory function with activation of T cell growth (3) augmentation of antigen-presenting function Key molecular interactions between T cells and APCs
 induction of cytokines/chemokines (IL-8, IL-12, TNF- , MIP-1 ) (2) stimulation of CD80 and CD86 expression and co- stimulatory function with activation of T cell growth (3) augmentation of antigen-presenting function Key molecular interactions between T cells and APCs")
37
Naïve CD4+ T cells differentiate into Th1 and Th2 subsets Resting CD4+ cell “pTh” Activated CD4+ cell IL-2 IL-2 IFN- TNF IL-4 IL-5 IL-6 IL-10 IFN- , IL-12 IL-4, IL-13 IL-4 IL-10 (–) IFN- (–) Th1 Cells Th2 Cells Antigen + APC
 IFN- (–) Th1 Cells Th2 Cells Antigen + APC")
38
IL-2 IFN- TNF IL-4 IL-5 IL-6 IL-10 IL-4 IL-10 (–) IFN- (–) Th1 Cells Th2 Cells Functions of Th1 subsets Activate macrophages/dendritic cells augment antigen presentation induce delayed type hypersensitivity (DTH) responses important in eradicating intracellular pathogens (TB, leprosy, listeria mediate Th1 diseases (ie; rheumatoid arthritis, multiple sclerosis and type I diabetes Help B cells and induce humoral immunity mediate allergic and immediate hypersensitivity responses involved in antibody mediated immune diseases like SLE and ITP Functions of Th2 subsets Functions of T helper subsets
 IFN- (–) Th1 Cells Th2 Cells Functions of Th1 subsets Activate macrophages/dendritic cells augment antigen presentation induce delayed type hypersensitivity (DTH) responses important in eradicating intracellular pathogens (TB, leprosy, listeria mediate Th1 diseases (ie; rheumatoid arthritis, multiple sclerosis and type I diabetes Help B cells and induce humoral immunity mediate allergic and immediate hypersensitivity responses involved in antibody mediated immune diseases like SLE and ITP Functions of Th2 subsets Functions of T helper subsets")
39
MHC Class ll ANTIGEN Internalization of antigen/Ig Antigenic peptides bind to MHC class II molecules B cell BCR (SmIg) Peptide Antigen binds specifically to BCR (surface membrane Ig), is internalized into vesicles and cleaved into peptides which displace and bind to MHC class II molecules. The peptide/MHC complex is then transported to the surface membrane. Antigen Processing and Presentation by B cells

40
CD3 TCR CD4 Fc receptor IL-12 IL-1 IL-6 IL-12 TNF TGF- MHC class II IFN- cytotoxic granules IL-2 Receptor IL-2 MHC II Macrophage Activated Macrophage Activated Th1 Cell T Cell- Macrophage Interactions CD4 Th1 Cell CD28 B7 (CD80) CD4 0L CD28 TCR CD4 CD80
 CD4 0L CD28 TCR CD4 CD80")
41
Maximum number of different types of HLA molecules expressed on the cell surface Class I (HLA-A) Class I (HLA-B) Class I (HLA-C) Class II (HLA-DR) Class II (HLA-DQ) Class II (HLA-DP) Total 22200062220006 Nucleated cells Antigen presenting cells 2 4 16 Each of these MHC molecules selects its own T cell repertoire that only recognizes peptides presented by that particular type of MHC molecule (actually more)
 Class I (HLA-B) Class I (HLA-C) Class II (HLA-DR) Class II (HLA-DQ) Class II (HLA-DP) Total Nucleated cells Antigen presenting cells Each of these MHC molecules selects its own T cell repertoire that only recognizes peptides presented by that particular type of MHC molecule (actually more)")
42
V. Cytokines you need to know IL-2 (big family e.g. IL-7 & IL-15) IL-4 (small family inc. IL-13) IL-6 (large family inc. G-CSF) IL-10 (growing family) IL-12 (small family inc. IL-23) IFN- IFN- (large family) IL-1 IL-18 LT- TNF- CD40L FasL TGF- (very large family) Chemokines (see Fig. 11.6) Inflammatory Non-inflammatory See Figs. 11.1 (p244), 11.2 (p245), 11.3 (p248) Tables 11-3 (p249), 11.4 (p264) in Abbas Innate Adaptive √√ √√ √ √√ √√ √ √√ √√ √ √√ √ √√ √√ Type I & II Cytokine Receptors (Hematopoietin R.) Toll (TLR) /IL-1 Receptors TNF Related Receptors TGF- Receptors Chemokine Receptors
 IL-4 (small family inc. IL-13) IL-6 (large family inc. G-CSF) IL-10 (growing family) IL-12 (small family inc. IL-23) IFN- IFN- (large family) IL-1 IL-18 LT- TNF- CD40L FasL TGF- (very large family) Chemokines (see Fig. 11.6) Inflammatory Non-inflammatory See Figs (p244), 11.2 (p245), 11.3 (p248) Tables 11-3 (p249), 11.4 (p264) in Abbas Innate Adaptive √√ √√ √ √√ √√ √ √√ √√ √ √√ √ √√ √√ Type I & II Cytokine Receptors (Hematopoietin R.) Toll (TLR) /IL-1 Receptors TNF Related Receptors TGF- Receptors Chemokine Receptors.")
43
V. Cytokines you need to know IL-2 (big family e.g. IL-7 & IL-15) IL-4 (small family inc. IL-13) IL-6 (large family inc. G-CSF) IL-10 (growing family) IL-12 (small family inc. IL-23) IFN- IFN- (large family) IL-1 IL-18 LT- TNF- CD40L FasL TGF- (very large family) Chemokines (see Fig. 11.6) Inflammatory Non-inflammatory See Figs. 11.1 (p244), 11.2 (p245), 11.3 (p248) Tables 11-3 (p249), 11.4 (p264) in Abbas Innate Adaptive √√ √√ √ √√ √√ √ √√ √√ √ √√ √ √√ √√ Type I & II Cytokine Receptors (Hematopoietin R.) Toll (TLR) /IL-1 Receptors TNF Related Receptors TGF- Receptors Chemokine Receptors
 IL-4 (small family inc. IL-13) IL-6 (large family inc. G-CSF) IL-10 (growing family) IL-12 (small family inc. IL-23) IFN- IFN- (large family) IL-1 IL-18 LT- TNF- CD40L FasL TGF- (very large family) Chemokines (see Fig. 11.6) Inflammatory Non-inflammatory See Figs (p244), 11.2 (p245), 11.3 (p248) Tables 11-3 (p249), 11.4 (p264) in Abbas Innate Adaptive √√ √√ √ √√ √√ √ √√ √√ √ √√ √ √√ √√ Type I & II Cytokine Receptors (Hematopoietin R.) Toll (TLR) /IL-1 Receptors TNF Related Receptors TGF- Receptors Chemokine Receptors.")
44
IL-2 activates T-cells in an autocrine manner

46
VIII. Chemokines Important chemoattractantsImportant chemoattractants Regulate steady state and inflammatory leukocyte trafficRegulate steady state and inflammatory leukocyte traffic Signal through G-protein coupled receptorsSignal through G-protein coupled receptors Therefore good drug targets for big PharmaTherefore good drug targets for big Pharma Two chemokine receptors serve as co-recpetors for HIV infection (CXCR4 and CCR5)Two chemokine receptors serve as co-recpetors for HIV infection (CXCR4 and CCR5)
Two chemokine receptors serve as co-recpetors for HIV infection (CXCR4 and CCR5).")
47
Question: How do viruses that don ’ t infect “ professional APCs ” such as dendritic cells elicit a primary immune response? After all, virally-infected cells don ’ t normally traffic to 2˚ lymphoid organs

48
PVR expressed on non-hematopoietic cells. Infection with Poliovirus Endocytosis of virus, nuclear entry, synthesis of viral proteins in cytosol. Presentation of viral peptides on MHC Class I to CD8+ cytotoxic T-cells Proliferation of cytotoxic T-cells (CTLs) Perforin/granzyme- mediated cell death “Classic” view of CTL response against virus-infected cells Cross-priming of exogenous antigens by dendritic cells PVR expressed on non-hematopoietic cells. Infection with Poliovirus Cytopathic changes; recognition and phagocytosis by dendritic cell Phagosome-to-cytosol protein export; ubiquitin-mediated proteolysis of viral proteins; Presentation of peptide via MHC Class I Perforin/granzyme- mediated cell death of DC; proliferation of CD8+ CTL; Killing of virus-infected epithelial cells by CTL
 Perforin/granzyme- mediated cell death Classic view of CTL response against virus-infected cells Cross-priming of exogenous antigens by dendritic cells PVR expressed on non-hematopoietic cells. Infection with Poliovirus Cytopathic changes; recognition and phagocytosis by dendritic cell Phagosome-to-cytosol protein export; ubiquitin-mediated proteolysis of viral proteins; Presentation of peptide via MHC Class I Perforin/granzyme- mediated cell death of DC; proliferation of CD8+ CTL; Killing of virus-infected epithelial cells by CTL.")
49
Why do NK Cells Fail to Recognize Healthy Cells?

50
1. For cytotoxic CD8 T-cells, ligation of the TCR by MHC I/peptide + co-stimulation results in release of granzymes and perforin and/or FasL, leading to apoptosis of the target cells. 2. Viruses evade host defense, in part, by down-regulating MHC Class I. Uninfected dendritic cells circumvent this by “cross-priming”: phagocytosis of virus-infected cell and presentation of “exogenous” viral antigens on MHC Class I. 3. The innate immune system has a rapid onset and recognizes molecular patterns in a non-clonal fashion. 4. NK cells lack TCRs, but instead express both activating and inhibitory (e.g., KIRs) receptors at their surfaces. The relative expression and ligation of these receptors determines the outcome (i.e., killing or not) of the NK effector response. 5. Innate immune B-cells (e.g., B-1 cells and marginal zone B cells) recognize carbohydrate antigens, secrete IgM, and are not long-lived. 6. Innate immune T-cells ( T-cells, and NK T cells) recognize non-peptide antigens in non-classical MHC-like molecules. They mediate cytotoxicity & rapid cytokine secretion. Summary
 receptors at their surfaces. The relative expression and ligation of these receptors determines the outcome (i.e., killing or not) of the NK effector response. 5. Innate immune B-cells (e.g., B-1 cells and marginal zone B cells) recognize carbohydrate antigens, secrete IgM, and are not long-lived. 6. Innate immune T-cells ( T-cells, and NK T cells) recognize non-peptide antigens in non-classical MHC-like molecules. They mediate cytotoxicity & rapid cytokine secretion. Summary.")
Similar presentations



 or antibody mediated immunity: The total immunological.>")

 Alternative Complement System Acquired (Adaptive) -B Lymphocytes.>")



>")

 2. Immunity to intracellular pathogens NK cells, control of Th1/Th2 responses 3.>")






>")

 & MHC Complexes-Antigen Presentation>")
 Prof. Dr. Zahid Shakoor MBBS, Ph D (London) College of Medicine King Saud University.>")
