Download presentation
Presentation is loading. Please wait.

1
T Cell Receptor & T Cell Development

2
Questions for the next 2 lectures:
How do you generate a diverse T cell population with functional TCR rearrangements? How do you generate a T cell population that is self-MHC restricted? How do you ensure that those diverse T cell receptors are not-self reactive? How do you coordinate lineage specification with MHC specificity and coreceptor expression? - vs. T cell - CD4 vs. CD8

3
a key regulator of the immune system
T lymphocyte: a key regulator of the immune system

4
T Lymphocytes Arise from stem cells resident in the bone marrow and migrate to the thymus which serves as an indoctrination center where thymocytes must learn to distinguish self from nonself Thymocytes that cannot make this induction are eliminated, those that can may further differentiate, mature, and graduate as T lymphocytes and enter the circulation

5
Origin, generation and differentiation of T cells

6
T Cell Development T cell progenitors migrate from bone marrow and seed thymus. T cell progenitors undergo differentiation to CD4, CD8 and NKT cells in thymus. Mature CD4 and CD8 T cells circulate between blood and lymphoid tissues until they meet antigens presented on dendritic cells in lymphoid tissues. T cells further undergo maturation to become functional memory or effector T cells in LT

7
Thymic involution: Human thymus is fully developed before birth and increases in size until puberty. It then progressively shrinks during adult life. Most thymectomized adults have no problem in T cell immunity because they have enough memory T cells in the periphery, and these T cells are long-lived. Figure 5-2

8
T Cell Functions Collectively, T cells display a number of diverse functions: - They often function to initiate, regulate, and fine-tune humoral immune response - They are effector cells responsible for various types of cell mediated immune responses like; DTH, contact sensitivity, transplantation immunity, and cytotoxicity

9
T Cell Surface Molecules
TCR: A very diverse heterodimer that lacks a cytoplasmic tail that would allow direct cytoplasmic signaling once TCR binds an epitope CD3 Complex: It is composed of a group of six invariant accessory molecules; one CD3 , one CD3 , two CD3ε, and an intracytoplasmic homodimer of ξ or CD247 chains Cytoplasmic signaling occurs through CD3 that noncovalently associate with TCR

10
TCR Complex Figure 3-6

11
The CD3 complex is essential for both cell surface expression of the TCR and for signal transduction once the TCR recognizes an antigen Unlike antibodies that can readily bind free antigen, a TCR cannot bind soluble antigens, but only enzymatically cleaved fragments of larger peptides presented as peptide MHC (pMHC) complexes
 complexes.")
12
CD4 or CD8: Most mature T cells express CD4 or CD8 molecules, they function as important co-receptors in association with the TCR By binding to invariant portions of the MHC I (CD8) or MHC II (CD4), they serve to increase the interaction of the MHC-bound antigenic fragment and the TCR
 or MHC II (CD4), they serve to increase the interaction of the MHC-bound antigenic fragment and the TCR.")
13
The structures of CD4 and CD8
Figure 3-10 13

14
CD8 binds MHC class I CD4 binds MHC class II
Most mature T cells are either CD4+ or CD8+. CD8 T cells kill cells infected with intracellular pathogens or tumor cells while CD4 T cells regulate (activate or suppress) other immune cells’ function. 14
 other immune cells’ function. 14.")
15
TCR Vs Immunoglobulin Both: Bind antigen
Have Variable region and Constant region Have a binding site that is a heterodimer (composed of 2 different chains) TCRs act only as receptors Igs act as receptors and effector molecules (soluble antigen-binding molecules)
 TCRs act only as receptors. Igs act as receptors and effector molecules. (soluble antigen-binding molecules)")
16
Similarity between TCR and Ig

17
TCR Structure The TCR is a member of the immunoglobulin supergene family and is composed of two polypeptide chains; a light α or chain and a heavy β or chain Each polypeptide chain of the heterodimer pair contains a variable and a constant region domain The Vα and V regions are encoded by V and J gene clusters The Vβ and V regions are encoded by V, D, and J gene clusters The D gene cluster provides an additional source of variation

18
Figure 3-7

19
a and b TCR gene loci (germline configuration)
")
20
The gene clusters undergo DNA rearrangement, similar to that already described for immunoglobulin genes, to synthesize αβ dimers or dimers As with immunoglobulins, the constant domain of the α and β or and chains are encode by constant region genes (Cα and Cβ or C or C ) T cell receptors do not undergo any subsequent changes equivalent to isotype switch, and somatic hypermutation, important to generating diversity of immunoglobulins.
 T cell receptors do not undergo any subsequent changes equivalent to isotype switch, and somatic hypermutation, important to generating diversity of immunoglobulins.")
21
As might be imagined, in the random process of generating diversity, a variety of TCR specificities would be generated for peptides that one may never encounter during his lifetime Three distinct categories of TCR specificities can be identified: - Those that recognize peptides that will never be encountered - Those that recognize peptides produced by potential pathogens or peptides of foreign origin - Those that recognize peptides that are produced by cells of self

22
TcR a gene rearrangement by SOMATIC RECOMBINATION
Germline TcR Vn J C V2 V1 Rearranged TcR 1° transcript Spliced TcR mRNA Rearrangement very similar to the IgL chains

23
TcR a gene rearrangement RESCUE PATHWAY
There is only a 1:3 chance of the join between the V and J region being in frame Vn J C V2 V1 Vn+1 chain tries for a second time to make a productive join using new V and J elements Productively rearranged TcR 1° transcript

24
TcR b gene rearrangement SOMATIC RECOMBINATION
L & V x52 D1 J C1 D2 C2 Germline TcR D-J Joining V-DJ joining Rearranged TcR 1° transcript C-VDJ joining Spliced TcR mRNA

25
TcR b gene rearrangement RESCUE PATHWAY
There is a 1:3 chance of productive D-J rearrangement and a 1:3 chance of productive V D-J rearrangement (i.e only a 1:9 chance of a productive b chain rearrangement) D1 J C1 D2 C2 Germline TcR D-J Joining V-DJ joining V 2nd chance at V-DJ joining Need to remove non productive rearrangement Use (DJC)b2 elements
 D1. J. C1. D2. C2. Germline TcR D-J Joining. V-DJ joining. V. 2nd chance at. V-DJ joining. Need to remove non productive. rearrangement. Use (DJC)b2 elements.")
26
TCR gene rearrangements occur in the thymus
The same RSS and the same enzymes are used to rearrange both the TCR genes and the Ig genes. P and N nucleotides are added at the junctions between rearranged segments n=70-80 n=52

27
TCR b Chain D region can be read in all frames
GTACTGCAGATT V J D J starts with ATT No additions: J is in frame GTACCTGCAGATT V J D One addition and J is out of frame; the ATT start for J is lost (as are all appropriate downstream codons) GTACCTGCAGGATT V J D Two addition and J is out of frame; the ATT start for J is lost (as are all appropriate downstream codons) GTACCTGCAGGCATT V J D Three addition and J is in frame; the ATT start for J is is present (as are all appropriate downstream codons)
 GTACCTGCAGGATT. V. J. D. Two addition and J is out of frame; the ATT start for J is lost (as are all appropriate downstream codons) GTACCTGCAGGCATT. V. J. D. Three addition and J is in frame; the ATT start for J is is present (as are all appropriate downstream codons)")
28
-chain locus is first to be rearranged
Wehfp wefnp ‘FFOEO

29
Two chances for productive (=correct reading frame) rearrangement: b chain
 rearrangement: b chain")
30
-chain rearrangement

31
Multiple rounds of -chain rearrangement can rescue nonproductive TCR
Wehfp wefnp ‘FFOEO

32
TCR gene rearrangement generates the TCR repertoire
Pre-TCR complex stops further gene rearrangement at b locus, and induces thymocyte proliferation Finally TCR+ DP cells are made 32

33
Germline configuration of g and d loci
TCRd D, J and C exons are encoded in the intron between the the Vs and the Js of the TCRa locus. The V segments for TCRd (4 known) are mixed in with the V segments of the TCRa
 are mixed in with the V segments of the TCRa.")
35
Figure 3-8 part 2 of 2 Most gd T cells do not express CD4 or CD8.
They are thought to be: First line of defense? Bridge between innate and adaptive responses? Figure 3-8 part 2 of 2

36
Signals through the TCR and the pre-TCR compete
to determine thymocyte lineage Wehfp wefnp ‘FFOEO 36

37
Generation of diversity in the TcR
COMBINATORIAL DIVERSITY Multiple germline segments In the human TcR Variable (V) segments: ~70, 52 Diversity (D) segments: 0, 2 Joining (J) segments: 61, 13 The need to pair and chains to form a binding site doubles the potential for diversity JUNCTIONAL DIVERSITY Addition of non-template encoded (N) and palindromic (P) nucleotides at imprecise joints made between V-D-J elements SOMATIC MUTATION IS NOT USED TO GENERATE DIVERSITY IN TcR
 segments: ~70, 52 Diversity (D) segments: 0, 2 Joining (J) segments: 61, 13 The need to pair and chains to form a binding site. doubles the potential for diversity. JUNCTIONAL DIVERSITY. Addition of non-template encoded (N) and palindromic (P) nucleotides at imprecise joints made between V-D-J elements. SOMATIC MUTATION IS NOT USED TO. GENERATE DIVERSITY IN TcR.")
38
The Generation of Diversity (GOD)
")
39
T Cell Development T cell precursors migrate from the bone marrow to enter the thymus as thymocytes, they express neither αβTCR nor CD4 or CD8 and are called double negative (DN) cells DN cells proliferate in the subcapsular region of the thymus and differentiate to express low levels of newly generated αβTCR, both CD4 and CD8, and are called double positive (DP) cells
 cells. DN cells proliferate in the subcapsular region of the thymus and differentiate to express low levels of newly generated αβTCR, both CD4 and CD8, and are called double positive (DP) cells.")
40
DP cells move inward to the deeper portion of the thymus, where they are fated to die within 3-4 days, unless their TCRs recognize an MHC class I or class II molecules on thymic dendritic cells. This process is called positive selection Although the mechanism of positive selection is yet unclear, partial recognition of class II by CD4 or class I by CD8 molecules must occur T cells that recognize self MHC molecules survive

41
A DP thymocyte with a TCR that engage MHC class I may become a CD8+ T cell and a DP thymocyte that recognizes MHC class II may become a CD4+ T cell Class I and class II molecules are not displayed on cell surface unless they are loaded with a peptide Only molecules of self origin are available on thymic APCs, and these are presented to the DP thymocyte in the deep or medullary area of the thymus

42
CD4+CD8+ DP cells: To be CD4 or CD8?
42

43
Thymocytes that show strong interaction with MHC molecules or pMHC complexes undergo apoptosis, a process known as negative selection Thymocytes that survive both positive and negative selection migrate from the thymus to populate lymphoid tissues and organs as T cells

44
Figure 5-3 part 2 of 2

45
Each thymocyte maturation stage occurs at
a distinct location of the thymus Wehfp wefnp ‘FFOEO Young adult:~5x107 thymocytes produced/day ~1.5x106 mature cells leave/day

46
Figure 5-3 part 1 of 2 DN (CD4-CD8-) and DP (CD4+CD8+) Immature
thymocytes are here Differentiation More mature SP (CD4+CD8-or CD8+CD4-) thymocytes are here
 thymocytes are here.")
47
Positive Selection Positive selection selects T cells that recognize peptides on self MHC This is to assure that mature T cells can respond to antigen-presented on self MHC. Self MHC I and II harboring self peptides on thymic epithelial cells recognize and activate TCRs on some DP thymocytes. DP thymocytes should receive this signal within 3-4 days to survive, otherwise they undergo apoptosis.

48
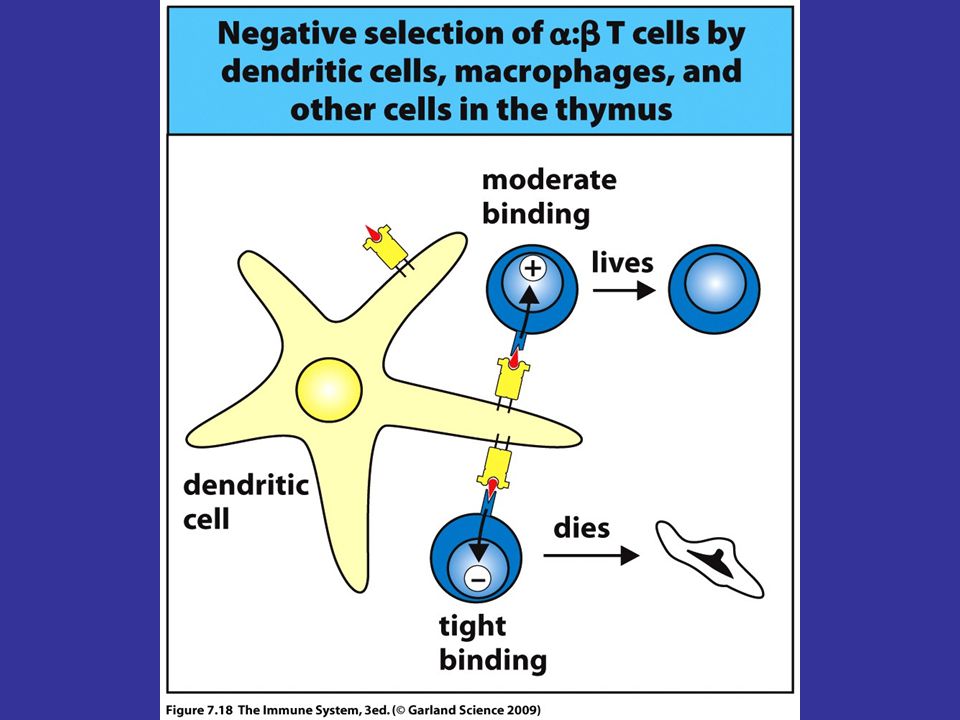
Negative Selection Negative selection eliminates T cells with TCRs that bind too strongly to self antigen/MHC complex (autoreactive cells are removed by this process) Dendritic cells and macrophages in cortico-medullary junction mediate it. Negative selection cannot eliminate T cells whose receptors are specific for self peptides that are present outside of the thymus These cells enter circulation, but soon to be rendered anergic or unresponsive by other mechanims.
 Dendritic cells and macrophages in cortico-medullary junction mediate it. Negative selection cannot eliminate T cells whose receptors are specific for self peptides that are present outside of the thymus. These cells enter circulation, but soon to be rendered anergic or unresponsive by other mechanims.")
50
Does receptor occupancy explain positive and negative selection ?
Low occupancy Survival High occupancy Negative selection

51
Does the TIME of receptor occupancy explain
positive and negative selection ? Long occupancy Negative selection Short occupancy Survival Short signaling Long signaling

52
Stage of maturation can be distinguished
by the expression of specific surface molecules DN DP Wehfp wefnp ‘FFOEO SP

53
Types of T cells Conventional: Uses ab TCR
Helper (CD4+) and cytotoxic (CD8+) T cells More abundant and highly specific Restricted by classical MHC (I and II) molecules Non-conventional: Uses gd TCR Primitive with broad specificity Restricted by non-classical molecules
 and cytotoxic (CD8+) T cells. More abundant and highly specific. Restricted by classical MHC (I and II) molecules. Non-conventional: Uses gd TCR. Primitive with broad specificity. Restricted by non-classical molecules.")
54
CD4+ T cells T cells with CD4 marker (glycoprotein) represent 70% of T cells in the periphery Named T helper cells Play central role in modulating cellular immunity via secretion of cytokines that mediate: B cell activation Immunoglobulin secretion (quality) Macrophage and dendritic cell activation Cellular chemotaxis and inflammation Two subsets; Th1 and Th2 cells
 Macrophage and dendritic cell activation. Cellular chemotaxis and inflammation. Two subsets; Th1 and Th2 cells.")
55
Th1 and Th2 cells CD4+ T helper cells can be classified into two types based on their cytokine profiles: T helper cell type 1 (Th1) and T helper cell type 2 (Th2). Cytokine profile is influenced by several factors: Nature and dose of antigen Route of administration Type of antigen presenting cell/ costimulation Genetic background The cytokine profile determines the effector function of the helper cell
 and T helper cell type 2 (Th2). Cytokine profile is influenced by several factors: Nature and dose of antigen. Route of administration. Type of antigen presenting cell/ costimulation. Genetic background. The cytokine profile determines the effector function of the helper cell.")
56
Differentiation of naive CD4 T cells into different subclasses
The differentiation of naive CD4 T cells into different subclasses of armed effector T cells is influenced by cytokines elicited by the pathogen. Many pathogens, especially intracellular bacteria and viruses, activate dendritic cells and NK cells to produce IL-12 and IFN-g, which cause proliferating CD4 T cells to differentiate into TH1 cells. IL-4 can inhibit these responses. IL-4, produced by an NK 1.1+ T cell in response to parasitic worms or other pathogens, acts on proliferating CD4 T cells to cause them to become TH2 cells. The mechanisms by which these cytokines induce the selective differentiation of CD4 T cells is now the subject of intensive study. They may act either when the CD4 T cell is first activated by an antigen-presenting cell or during the subsequent proliferative phase.

57
The nature and amount of ligand determine CD4 T cell functional phenotype
The nature and amount of ligand presented to a CD4 T cell during primary stimulation can determine its functional phenotype. CD4 T cells presented with low levels of a ligand that binds the T-cell receptor poorly differentiate preferentially into TH2 cells making IL-4 and IL-5. Such T cells are most active in stimulating naive B cells to differentiate into plasma cells and make antibody. T cells presented with a high density of a ligand that binds the T- cell receptor strongly differentiate into TH1 cells that secrete IL-2, TNF-b, and IFN-g, and are most effective in activating macrophages.

58
T Helper (CD4+ ) Subsets Antigen APC IL-4 IL-12 FasL Suicide Fas DR4
Anti-Inflammatory Cytokines Pro-Inflammatory Cytokines IL-4 IL-10 IL-13 IL-5 IL-6 IFN-g IL-2 LT Th1 response Cellular Immunity DTH Th2 response, Humoral Immunity and Acute Hypersensitivity

59
Differences between Th1 and Th2 cells
Produces type 1 cytokines IL-2, IFN- , TNF-α, TNF-β Activates macrophages and DCs for intracellular killing of pathogens Mediates CMI Produces type 2 cytokines IL-4, IL-5, IL-10, IL-13 Provides help to B cells in antibody response Mediates allergy and immunity to extracellular pathogens, including parasites

60
Cytotoxic T cells T cells that express CD8 molecule on their surface and they represent 30% of T cells in the periphery Destroy cells infected by intracellular pathogens and cancer cells Class I MHC molecules (nucleated body cells) expose foreign proteins TC cell releases perforin and granzymes, proteins that form pores in the target cell membrane; causing cell lysis and/or apoptosis
 expose foreign proteins. TC cell releases perforin and granzymes, proteins that form pores in the target cell membrane; causing cell lysis and/or apoptosis.")
62
Effector molecules of T cell subsets
The three main types of armed effector T cell produce distinct sets of effector molecules. CD8 T cells are predominantly killer T cells that recognize pathogen-derived peptides bound to MHC class I molecules. They release perforin (which creates holes in the target cell membrane), granzymes (which are proteases that act intracellularly to trigger apoptosis), and often the cytokine IFN-g. A membrane-bound effector molecule expressed on CD8 T cells is Fas ligand. When this binds to Fas on a target cell it activates apoptosis in the Fas-bearing cell. CD4 T cells recognize peptides bound to MHC class II molecules and are of two functional types: TH1 and TH2. TH1 cells are specialized to activate macrophages that are infected by or have ingested pathogens; they secrete IFN-g as well as other effector molecules, and express membrane-bound CD40 ligand and/or Fas ligand. These are both members of the TNF family but CD40 ligand triggers the activation of the target cell, whereas Fas ligand triggers the death of Fas-expressing cells, so their pattern of expression has a strong influence on their function. TH2 cells are specialized for B- cell activation; they secrete the B-cell growth factors IL-4 and IL-5. The principal membrane-bound effector molecule expressed by TH2 cells is CD40 ligand, which binds to CD40 on the B cell and induces B-cell proliferation.
, granzymes (which are proteases that act intracellularly to trigger apoptosis), and often the cytokine IFN-g. A membrane-bound effector molecule expressed on CD8 T cells is Fas ligand. When this binds to Fas on a target cell it activates apoptosis in the Fas-bearing cell. CD4 T cells recognize peptides bound to MHC class II molecules and are of two functional types: TH1 and TH2. TH1 cells are specialized to activate macrophages that are infected by or have ingested pathogens; they secrete IFN-g as well as other effector molecules, and express membrane-bound CD40 ligand and/or Fas ligand. These are both members of the TNF family but CD40 ligand triggers the activation of the target cell, whereas Fas ligand triggers the death of Fas-expressing cells, so their pattern of expression has a strong influence on their function. TH2 cells are specialized for B- cell activation; they secrete the B-cell growth factors IL-4 and IL-5. The principal membrane-bound effector molecule expressed by TH2 cells is CD40 ligand, which binds to CD40 on the B cell and induces B-cell proliferation.")
63
Regulation of the Immune Response
How does the immune system prevent self reactivity while maintaining reactivity to invaders/non-self? Clonal deletion/inactivation of auto-reactive cells Regulatory T cells keep potentially pathogenic self reactive T cells in check through “suppressive” mechanisms

64
New T cell phenotypes Regulatory T cells
Naturally occurring (CD25 positive) Induced (IL-10 and TGF-β) Some NK T cells Suppressor T cells Th17 cells: produce IL-17, role in acute inflammation, suppress Th2.
 Induced (IL-10 and TGF-β) Some NK T cells. Suppressor T cells. Th17 cells: produce IL-17, role in acute inflammation, suppress Th2.")
65
Bettelli et al., Nature 2008

66
Th17 and Treg

67
Regulatory T Cells (Treg)
CD3+, TcR+, CD4+, CD25+ Function Suppress the activity of effector Th and Tc cells Inhibition is antigen specific and MHC restricted Inhibition depends on cell:cell contact Appear to function in autoimmunity Development Develop in the bone marrow from DP cells Arise as a result of strong binding to self MHC and self Ag during negative selection. Alternative to cell death or anergy Development and maintenance are dependent upon B7

68
What are the requirements for Treg development?
TCR engagement in the thymus (high affinity but not so high as negative selection) Higher percentage of thymocytes with high affinity TCR in context of auto antigen develop into regs T cells of appropriate affinity are “instructed” to become Tregs Selective sparing of pre-committed cells from negative selection or promotion of Treg lineage development? higher percentage, but not higher absolute number Preferential elimination of non-regulatory T cells rather than increased production of Tregs TCR engagement serves as survival or expansion signal of Tregs pre-committed to that lineage.
 Higher percentage of thymocytes with high affinity TCR in context of auto antigen develop into regs. T cells of appropriate affinity are instructed to become Tregs. Selective sparing of pre-committed cells from negative selection or promotion of Treg lineage development higher percentage, but not higher absolute number. Preferential elimination of non-regulatory T cells rather than increased production of Tregs. TCR engagement serves as survival or expansion signal of Tregs pre-committed to that lineage.")
69
The Treg cell phenotype
CD4 Co-receptor for TCR recognition of MHC II/Ag CD25 IL-2Ra IL-2R component, confers high affinity binding to IL-2Rbg Key TR growth factor CTLA-4 cytotoxic T lymphocyte Ag-4 Binds to B7s (CD80/86) on APC, acts as co-stimulatory molecule for TR (blocking CTLA-4 inhibits TR) GITR glucocorticoid induced TNF related protein Ligation inhibits TR function (agonist inhibit TR, blocking augments TR) FoxP3 Forkhead /winged-helix TF critical for TR activity and development Unlike surface markers / receptors, TE do not express FoxP3 CD4 GITR CD25 CTLA-4 Treg Foxp3 The Treg cell phenotype
 on APC, acts as co-stimulatory molecule for TR (blocking CTLA-4 inhibits TR) GITR glucocorticoid induced TNF related. protein. Ligation inhibits TR function (agonist inhibit TR, blocking augments TR) FoxP3. Forkhead /winged-helix TF critical for TR activity and development. Unlike surface markers / receptors, TE do not express FoxP3. CD4. GITR. CD25. CTLA-4. Treg. Foxp3. The Treg cell phenotype.")
70
Modulation of immune responses by Treg cells
Treg cells are crucial for the induction and maintenance of peripheral tolerance to self-antigens Treg cells can also suppress immune responses to Tumor antigens Alloantigen Allergens Microbial antigens Teff Treg Tumor Clearance Microbial Immunity Autoimmunity Transplant rejection Graft-versus host disease Allergy Teff Treg Self-tolerance Transplantation tolerance Tumor Progression Microbial Persistence Teff Treg Self-tolerance Immunocompetence Sheng Cai

Similar presentations



















>")

